
Abstract
Anaerobic methane oxidation is coupled to the reduction of electron acceptors, such as sulfate, and contributes to their biogeochemical cycling in the environment. However, whether arsenate acts as an alternative electron acceptor of anaerobic methane oxidation and how this influences global arsenic transformations remains elusive. Here, we present incubations of arsenate-contaminated wetland soils from seven provinces in China. Using isotopically labelled methane, we find that anaerobic methane oxidation was linked to arsenate reduction at a rate approaching the theoretical arsenic/methane stoichiometric ratio of 4. In microcosm incubations with natural wetland soils, we find that the coupled pathway of anaerobic methane oxidation and arsenate reduction contributed 26 to 49% of total arsenic release from soils, with arsenic in the more soluble and toxic form arsenite. Comparative gene quantification and metagenomic sequencing suggest that the coupled pathway was facilitated by anaerobic methanotrophs, either independently or synergistically with arsenate-reducing bacteria through reverse methanogenesis and respiratory arsenate reduction. Further bioinformatic analyses show that genes coding for reverse methanogenesis and respiratory arsenate reduction are universally co-distributed in nature. This suggests that coupling of anaerobic methane oxidation and arsenate reduction is a potentially global but previously overlooked process, with implications for arsenic mobilization and environmental contamination.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$259.00 per year
only $21.58 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout






Similar content being viewed by others


Data availability
All the metagenomic sequencing data have been submitted to the Sequence Read Archive, with BioProject accession number of PRJNA612595. The source data have been deposited into the Open Science Framework (OSF) at https://doi.org/10.17605/OSF.IO/N4SEF. Source data are provided with this paper.
References
Zhu, Y. G., Yoshinaga, M., Zhao, F. J. & Rosen, B. P. Earth abides arsenic biotransformations. Annu. Rev. Earth Planet. Sci. 42, 443–467 (2014).
Singh, R., Singh, S., Parihar, P., Singh, V. P. & Prasad, S. M. Arsenic contamination, consequences and remediation techniques: a review. Ecotoxicol. Environ. Saf. 112, 247–270 (2015).
Cullen, W. R. & Reimer, K. J. Arsenic speciation in the environment. Chem. Rev. 89, 713–764 (1989).
Masscheleyn, P. H., Delaune, R. D. & Patrick, W. H. Effect of redox potential and pH on arsenic speciation and solubility in a contaminated soil. Environ. Sci. Technol. 25, 1414–1419 (1991).
Abernathy, C. O., Thomas, D. J. & Calderon, R. L. Health effects and risk assessment of arsenic. J. Nutr. 133, 1536S–1538S (2003).
Jia, Y. et al. Microbial arsenic methylation in soil and rice rhizosphere. Environ. Sci. Technol. 47, 3141–3148 (2013).
Chen, C. et al. Sulfate-reducing bacteria and methanogens are involved in arsenic methylation and demethylation in paddy soils. ISME J. 13, 2523–2535 (2019).
Burton, E. D., Johnston, S. G. & Kocar, B. D. Arsenic mobility during flooding of contaminated soil: the effect of microbial sulfate reduction. Environ. Sci. Technol. 48, 13660–13667 (2014).
Bao, P. et al. The role of sulfate-reducing prokaryotes in the coupling of element biogeochemical cycling. Sci. Total Environ. 613, 398–408 (2018).
Wolfe, A. L. & Wilkin, R. T. Evidence of sulfate-dependent anaerobic methane oxidation within an area impacted by coalbed methane-related gas migration. Environ. Sci. Technol. 51, 1901–1909 (2017).
Amstaetter, K., Borch, T., Larese-Casanova, P. & Kappler, A. Redox transformation of arsenic by Fe(II)-activated goethite (alpha-FeOOH). Environ. Sci. Technol. 44, 102–108 (2010).
Ma, J. F. et al. Transporters of arsenite in rice and their role in arsenic accumulation in rice grain. Proc. Natl Acad. Sci. USA 105, 9931–9935 (2008).
Zobrist, J., Dowdle, P. R., Davis, J. A. & Oremland, R. S. Mobilization of arsenite by dissimilatory reduction of adsorbed arsenate. Environ. Sci. Technol. 34, 4747–4753 (2000).
Macur, R. E., Wheeler, J. T., McDermott, T. R. & Inskeep, W. P. Microbial populations associated with the reduction and enhanced mobilization of arsenic in mine tailings. Environ. Sci. Technol. 35, 3676–3682 (2001).
Bruschi, M., Barton, L. L., Goulhen, F. & Plunkett, R. M. in Sulphate-Reducing Bacteria: Environmental and Engineered Systems (eds Barton, L. L. & Hamilton, W. A.) 435–458 (Cambridge Univ. Press, 2007).
Qiao, J. T. et al. Transcriptional activity of arsenic-reducing bacteria and genes regulated by lactate and biochar during arsenic transformation in flooded paddy soil. Environ. Sci. Technol. 52, 61–70 (2018).
Kudo, K. et al. Release of arsenic from soil by a novel dissimilatory arsenate-reducing bacterium, Anaeromyxobacter sp. strain PSR-1. Appl. Environ. Microbiol. 79, 4635–4642 (2013).
Qiao, J. et al. Humic substances facilitate arsenic reduction and release in flooded paddy soil. Environ. Sci. Technol. 53, 5034–5042 (2019).
Milucka, J. et al. Zero-valent sulphur is a key intermediate in marine methane oxidation. Nature 491, 541–546 (2012).
Raghoebarsing, A. A. et al. A microbial consortium couples anaerobic methane oxidation to denitrification. Nature 440, 918–921 (2006).
Beal, E. J., House, C. H. & Orphan, V. J. Manganese- and iron-dependent marine methane oxidation. Science 325, 184–187 (2009).
Devol, A. H. & Ahmed, S. I. Are high rates of sulfate reduction associated with anaerobic oxidation of methane. Nature 291, 407–408 (1981).
Niemann, H. et al. Novel microbial communities of the Haakon Mosby mud volcano and their role as a methane sink. Nature 443, 854–858 (2006).
Boetius, A. et al. A marine microbial consortium apparently mediating anaerobic oxidation of methane. Nature 407, 623–626 (2000).
Orphan, V. J., House, C. H., Hinrichs, K. U., McKeegan, K. D. & DeLong, E. F. Methane-consuming archaea revealed by directly coupled isotopic and phylogenetic analysis. Science 293, 484–487 (2001).
Haroon, M. F. et al. Anaerobic oxidation of methane coupled to nitrate reduction in a novel archaeal lineage. Nature 500, 567–570 (2013).
Cai, C. et al. A methanotrophic archaeon couples anaerobic oxidation of methane to Fe(III) reduction. ISME J. 12, 1929–1939 (2018).
Leu, A. O. et al. Anaerobic methane oxidation coupled to manganese reduction by members of the Methanoperedenaceae. ISME J. 14, 1030–1041 (2020).
Caldwell, S. L. et al. Anaerobic oxidation of methane: mechanisms, bioenergetics, and the ecology of associated microorganisms. Environ. Sci. Technol. 42, 6791–6799 (2008).
Leu, A. O. et al. Lateral gene transfer drives metabolic flexibility in the anaerobic methane-oxidizing archaeal family Methanoperedenaceae. mBio 11, e01325-20 (2020).
Ruff, S. E. et al. Methane seep in shallow-water permeable sediment harbors high diversity of anaerobic methanotrophic communities, Elba, Italy. Front. Microbiol. 7, 374 (2016).
Hu, B. L. et al. Evidence for nitrite-dependent anaerobic methane oxidation as a previously overlooked microbial methane sink in wetlands. Proc. Natl Acad. Sci. USA 111, 4495–4500 (2014).
Muehe, E. M., Wang, T. M., Kerl, C. F., Planer-Friedrich, B. & Fendorf, S. Rice production threatened by coupled stresses of climate and soil arsenic. Nat. Commun. 10, 4985 (2019).
Seyfferth, A. L., McCurdy, S., Schaefer, M. V. & Fendorf, S. Arsenic concentrations in paddy soil and rice and health implications for major rice-growing regions of Cambodia. Environ. Sci. Technol. 48, 4699–4706 (2014).
Sohn, E. The toxic side of rice. Nature 514, S62–S63 (2014).
Das, S., Liu, C. C., Jean, J. S. & Liu, T. L. Dissimilatory arsenate reduction and in situ microbial activities and diversity in arsenic-rich groundwater of Chianan Plain, Southwestern Taiwan. Microbiol. Ecol. 71, 365–374 (2016).
Jiang, J., Bauer, I., Paul, A. & Kappler, A. Arsenic redox changes by microbially and chemically formed semiquinone radicals and hydroquinones in a humic substance model quinone. Environ. Sci. Technol. 43, 3639–3645 (2009).
Shen, L. D., Ouyang, L., Zhu, Y. & Trimmer, M. Active pathways of anaerobic methane oxidation across contrasting riverbeds. ISME J. 13, 752–766 (2019).
Aromokeye, D. A. et al. Rates and microbial players of iron-driven anaerobic oxidation of methane in methanic marine sediments. Front. Microbiol. 10, 3041 (2020).
Wang, F. P. et al. Methanotrophic archaea possessing diverging methane-oxidizing and electron-transporting pathways. ISME J. 8, 1069–1078 (2014).
Hallam, S. J. et al. Reverse methanogenesis: testing the hypothesis with environmental genomics. Science 305, 1457–1462 (2004).
Wegener, G., Krukenberg, V., Riedel, D., Tegetmeyer, H. E. & Boetius, A. Intercellular wiring enables electron transfer between methanotrophic archaea and bacteria. Nature 526, 587–590 (2015).
McGlynn, S. E., Chadwick, G. L., Kempes, C. P. & Orphan, V. J. Single cell activity reveals direct electron transfer in methanotrophic consortia. Nature 526, 531–535 (2015).
Scheller, S., Yu, H., Chadwick, G. L., McGlynn, S. E. & Orphan, V. J. Artificial electron acceptors decouple archaeal methane oxidation from sulfate reduction. Science 351, 703–707 (2016).
Kletzin, A. et al. Cytochromes c in Archaea: distribution, maturation, cell architecture, and the special case of Ignicoccus hospitalis. Front. Microbiol. 6, 439 (2015).
Zhu, Y. G., Xue, X. M., Kappler, A., Rosen, B. P. & Meharg, A. A. Linking genes to microbial biogeochemical cycling: lessons from arsenic. Environ. Sci. Technol. 51, 7326–7339 (2017).
Shi, L. D. et al. Multi-omics reveal various potential antimonate reductases from phylogenetically diverse microorganisms. Appl. Microbiol. Biotechnol. 103, 9119–9129 (2019).
Ahmann, D., Roberts, A. L., Krumholz, L. R. & Morel, F. M. M. Microbe grows by reducing arsenic. Nature 371, 750 (1994).
Tian, H. X., Shi, Q. T. & Jing, C. Y. Arsenic biotransformation in solid waste residue: comparison of contributions from bacteria with arsenate and iron reducing pathways. Environ. Sci. Technol. 49, 2140–2146 (2015).
Shi, L. et al. Extracellular electron transfer mechanisms between microorganisms and minerals. Nat. Rev. Microbiol. 14, 651–662 (2016).
Lovley, D. R. & Malvankar, N. S. Seeing is believing: novel imaging techniques help clarify microbial nanowire structure and function. Environ. Microbiol. 17, 2209–2215 (2015).
Zhao, F. J., Ma, J. F., Meharg, A. A. & McGrath, S. P. Arsenic uptake and metabolism in plants. New Phytol. 181, 777–794 (2009).
Bao, S. D. Soil Agrochemical Analysis 3rd edn (China Agricultural Press, 2011).
Teske, A. et al. Microbial diversity of hydrothermal sediments in the Guaymas Basin: evidence for anaerobic methanotrophic communities. Appl. Environ. Microbiol. 68, 1994–2007 (2002).
Biddle, J. F. et al. Anaerobic oxidation of methane at different temperature regimes in Guaymas Basin hydrothermal sediments. ISME J. 6, 1018–1031 (2012).
Oswald, K. et al. Aerobic gammaproteobacterial methanotrophs mitigate methane emissions from oxic and anoxic lake waters. Limnol. Oceanogr. 61, S101–S118 (2016).
Nunoura, T. et al. Quantification of mcrA by quantitative fluorescent PCR in sediments from methane seep of the Nankai Trough. FEMS Microbiol. Ecol. 57, 149–157 (2006).
Vaksmaa, A., Jetten, M. S. M., Ettwig, K. F. & Lüke, C. McrA primers for the detection and quantification of the anaerobic archaeal methanotroph ‘Candidatus Methanoperedens nitroreducens’. Appl. Microbiol. Biotechnol. 101, 1631–1641 (2017).
Song, B., Chyun, E., Jaffe, P. R. & Ward, B. B. Molecular methods to detect and monitor dissimilatory arsenate-respiring bacteria (DARB) in sediments. FEMS Microbiol. Ecol. 68, 108–117 (2009).
Paszczynski, A. J. et al. Proteomic and targeted qPCR analyses of subsurface microbial communities for presence of methane monooxygenase. Biodegradation 22, 1045–1059 (2011).
Zhao, H. P. et al. Interactions between perchlorate and nitrate reductions in the biofilm of a hydrogen-based membrane biofilm reactor. Environ. Sci. Technol. 45, 10155–10162 (2011).
Freitag, T. E. & Prosser, J. I. Correlation of methane production and functional gene transcriptional activity in a peat soil. Appl. Environ. Microbiol. 75, 6679–6687 (2009).
Bolger, A. M., Lohse, M. & Usadel, B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120 (2014).
Li, D. et al. MEGAHIT v1.0: a fast and scalable metagenome assembler driven by advanced methodologies and community practices. Methods 102, 3–11 (2016).
Dong, X. L. & Strous, M. An integrated pipeline for annotation and visualization of metagenomic contigs. Front. Genet. 10, 999 (2019).
Eddy, S. R. Accelerated profile HMM searches. PLoS Comput. Biol. 7, e1002195 (2011).
Saunders, J. K., Fuchsman, C. A., McKay, C. & Rocap, G. Complete arsenic-based respiratory cycle in the marine microbial communities of pelagic oxygen-deficient zones. Proc. Natl Acad. Sci. USA 116, 9925–9930 (2019).
Price, M. N., Dehal, P. S. & Arkin, A. P. FastTree 2—approximately maximum-likelihood trees for large alignments. PLoS ONE 5, e9490 (2010).
Edgar, R. C. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 32, 1792–1797 (2004).
Ma, L. P. et al. Metagenomic assembly reveals hosts of antibiotic resistance genes and the shared resistome in pig, chicken, and human feces. Environ. Sci. Technol. 50, 420–427 (2016).
Finn, R. D. et al. Pfam: the protein families database. Nucleic Acids Res. 42, D222–D230 (2014).
Water Quality – Determination of Iron-phenanthroline Spectrophotometry HJ/T 345-2007 (in Chinese) (Ministry of Ecology and Environment of the People’s Republic of China, 2007).
China Climate Bulletin in 2019 (in Chinese) (China Meteorological Administration, 2020).
Mitchell, A. L. et al. MGnify: the microbiome analysis resource in 2020. Nucleic Acids Res. 48, D570–D578 (2019).
Buchfink, B., Xie, C. & Huson, D. H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 12, 59–60 (2015).
Acknowledgements
The authors greatly thank the National Natural Science Foundation of China (grants 51878596, 41991332 and 21577123), the Natural Science Funds for Distinguished Young Scholar of Zhejiang Province (LR17B070001) and the National Key Technology R&D Program (2018YFC1802203) for their financial support.
Author information
Authors and Affiliations
Contributions
L.-D.S. conceived and conducted the incubations with soil inocula, performed and analysed metagenomics and other bioinformatics, evaluated and arranged the results, and drafted the manuscript; T.G. took natural samples, measured physicochemical characteristics and quantified the functional genes/transcripts; P.-L.L. and Z.-F.N. helped perform the experiments inoculated by soil samples and microcosm incubations; Y.-J.Z. performed the microcosm incubations; X.-J.T. supervised the project, conceived the experiments and wrote the manuscript; P.Z., L.-Z.Z., Y.-G.Z. and A.K. assisted in design and set-up of the project and contributed to the manuscript preparation; H.-P.Z. initiated and supervised the project, conceived the experiments and wrote the manuscript; all authors contributed to revising the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Peer review information Primary Handling Editors: Clare Davis; Xujia Jiang.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
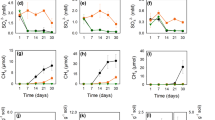
Extended Data Fig. 1 Reduced As(V) and produced 13DIC of experiments inoculated by soil samples during the whole incubation (7 days).
The main differences between these two groups are 13CH4 (top comparison) and As(V) (bottom comparison), respectively. Error bars indicate standard deviations of triplicate setups. The value of ΔAs(III)/ΔAs(V) represents the ratio of increased dissolved As(III) to decreased dissolved As(V). Asterisks indicate the statistical significance of the difference between experiment setups and corresponding control setups calculated by Kruskal-Wallis test.
Extended Data Fig. 2 Stoichiometry and kinetics of AOM-AsR for the AH-MAS-1 soil sample.
Mass balance was tested for 13CH4 (a) and As(V) (b). Electron balance between 13CH4 oxidation and As(V) reduction was studied in terms of rate (c) and ratio (d). Error bars indicate standard deviations of triplicate setups. The confidence levels for fitting curves are 95%. Please note the different y-axis scales for different items.
Extended Data Fig. 3 Gene abundance and correlation with methane oxidation and arsenate reduction rates.
Absolute abundances of mcrA and arrA (copies/g dry soil) were normalized by logarithm with a base of ten (a). Gene abundances and reaction rates were correlated using Spearman rank correlation (b and c), where ρ and p indicate the correlation efficiency and statistic probability, respectively. Error bars indicate standard deviations of triplicates. The points (SC-MY-1, YN-KM, and AH-MAS-1) in the grey circle (c) of which standard residual is 2-fold more than the standard deviation are removed based on Casewise Diagnostics. Their relatively high ATL values might explain why As(V) could be reduced rapidly at low arrA abundances (Supplementary Fig. 4).
Extended Data Fig. 4 Detailed phylogeny of respiratory arsenate reductase.
The proteins recovered in this study are highlighted in purple. Bootstrap values are generated from 100 replicates. The scale bar represents amino acid changes.
Extended Data Fig. 5 Phylogeny of respiratory arsenate reductase beta subunit (ArrB) and closely related anaerobic arsenite oxidase beta subunit (ArxB).
The proteins recovered in this study are highlighted in purple. A 4Fe-4S ferredoxin is used as the outgroup. Bootstrap values are generated from 100 replicates. The scale bar represents amino acid changes.
Extended Data Fig. 6 Putative metabolic pathways of AOM-AsR.
Methanoperedenaceae members perform AOM-AsR independently using arsenate reductase acquired through lateral gene transfer (a). Electrons are generated from methane oxidation via the reverse methanogenesis pathway in ANME groups, and then transferred to arsenate reducers possibly through multi-haem cytochromes (b). Abbreviations: Mcr, methyl-coenzyme M reductase; MHC, multi-haem cytochrome; Arr, respiratory arsenate reductase.
Extended Data Fig. 7 Contents of 13CO2 in the headspace in the microcosm incubations.
(a), (c), and (e) are incubated with YN-MG-2, SC-CD-1, and SC-MY-2, respectively, while (b), (d), and (f) use the corresponding autoclaved samples. The dark and light grey in each figure indicate the experimental setups amended with environmentally relevant concentrations of 13CH4, and the control setups without methane addition, respectively. The probability (i.e. p-values) of the differences between two setups was calculated using Kruskal-Wallis test. Error bars indicate standard deviations of triplicate setups.
Extended Data Fig. 8 Concentrations of As(V) and As(III) in the porewaters in the microcosm incubations.
(a), (c), and (e) were incubated with YN-MG-2, SC-CD-1, and SC-MY-2, respectively, while (b), (d), and (f) show the corresponding autoclaved samples. The dark and light red in each figure indicate the experimental setups amended with environmentally relevant concentrations of 13CH4, and the control setups without methane addition, respectively. The upper parts show As(III) while the bottom ones show As(V). Please note the different y-axis scales in the different panels. The probability (i.e. p-values) of differences between two setups was calculated using Kruskal-Wallis test. Error bars indicate standard deviations of triplicate setups.
Extended Data Fig. 9 Concentrations of total Fe and Fe(II) in the porewaters in the microcosm incubations.
(a), (c), and (e) were incubated with YN-MG-2, SC-CD-1, and SC-MY-2, respectively, while (b), (d), and (f) used the corresponding autoclaved samples. The dark and light pink in each figure indicate the experimental setups amended with environmentally relevant concentrations of 13CH4, and the control setups without methane addition, respectively. The upper parts show total Fe while the bottom ones show Fe(II). Please note the different y-axis scales in the different panels. The probability (i.e. p-values) of differences between two setups was calculated using Kruskal-Wallis test. Error bars indicate standard deviations of triplicate setups.
Extended Data Fig. 10 Concentrations of total Mn in the porewaters during the microcosm incubations.
(a), (c), and (e) were incubated with YN-MG-2, SC-CD-1, and SC-MY-2, respectively, while (b), (d), and (f) used the corresponding autoclaved samples. The dark and light green in each figure indicate the experimental setups amended with environmentally relevant concentrations of 13CH4, and the control setups without methane addition, respectively. The probability (i.e. p-values) of differences between two setups was calculated using Kruskal-Wallis test. Error bars indicate standard deviations of triplicate setups.
Supplementary information
Supplementary Information
Supplementary Discussion, Figs. 1–11, Tables 1–4, captions for Supplementary Datasets 1–3 and references for Supplementary Information citations.
Supplementary Dataset 1
Geographical information of collected natural soil samples.
Supplementary Dataset 2
Physicochemical characteristics of collected natural soil samples.
Supplementary Dataset 3
Geographical information of bioinformatic predicted samples.
Source data
Source Data Fig. 2
Numerical data for Fig. 2.
Source Data Fig. 5
Numerical data for Fig. 5.
Rights and permissions
About this article

Cite this article
Shi, LD., Guo, T., Lv, PL. et al. Coupled anaerobic methane oxidation and reductive arsenic mobilization in wetland soils. Nat. Geosci. 13, 799–805 (2020). https://doi.org/10.1038/s41561-020-00659-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41561-020-00659-z
This article is cited by
-
Mechanisms of extracellular electron transfer in anaerobic methanotrophic archaea
Nature Communications (2024)
-
Unexpected genetic and microbial diversity for arsenic cycling in deep sea cold seep sediments
npj Biofilms and Microbiomes (2023)
-
Unraveling the role of sulfide-natural organic matter interplay on methane cycling in anoxic environments
Biogeochemistry (2022)
-
Biochar for agronomy, animal farming, anaerobic digestion, composting, water treatment, soil remediation, construction, energy storage, and carbon sequestration: a review
Environmental Chemistry Letters (2022)
-
Methane-dependent selenate reduction by a bacterial consortium
The ISME Journal (2021)
