
Abstract
Higher-order chromatin structure regulates gene expression, and mutations in proteins mediating genome folding underlie developmental disorders known as cohesinopathies. However, the relationship between three-dimensional genome organization and embryonic development remains unclear. Here we define a role for bromodomain-containing protein 4 (BRD4) in genome folding, and leverage it to understand the importance of genome folding in neural crest progenitor differentiation. Brd4 deletion in neural crest results in cohesinopathy-like phenotypes. BRD4 interacts with NIPBL, a cohesin agonist, and BRD4 depletion or loss of the BRD4–NIPBL interaction reduces NIPBL occupancy, suggesting that BRD4 stabilizes NIPBL on chromatin. Chromatin interaction mapping and imaging experiments demonstrate that BRD4 depletion results in compromised genome folding and loop extrusion. Finally, mutation of individual BRD4 amino acids that mediate an interaction with NIPBL impedes neural crest differentiation into smooth muscle. Remarkably, loss of WAPL, a cohesin antagonist, rescues attenuated smooth muscle differentiation resulting from BRD4 loss. Collectively, our data reveal that BRD4 choreographs genome folding and illustrates the relevance of balancing cohesin activity for progenitor differentiation.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout








Similar content being viewed by others


Data availability
All data generated for this paper have been deposited at the National Center for Biotechnology Information Gene Expression Omnibus (http://www.ncbi.nlm.nih.gov/geo) and are available under accession no. GSE169516.
Mouse genome sequence mm10:
Saccharomyces cerevisiae genome sequence sacCer3: https://hgdownload.soe.ucsc.edu/goldenPath/sacCer3/bigZips/sacCer3.fa.gz
BRD4 ChIP–seq: http://dc2.cistrome.org/api/hgtext/34084/?db=mm10
NIPBL ChIP–seq: http://dc2.cistrome.org/api/hgtext/2864/?db=mm10
H3K27ac ChIP–seq: http://dc2.cistrome.org/api/hgtext/67169/?db=mm10
RAD21 ChIP–seq: http://dc2.cistrome.org/api/hgtext/39300/?db=mm10.Source data are provided with this paper.
Code availability
OligoMiner is available at https://github.com/beliveau-lab/OligoMiner and cLoops at https://github.com/YaqiangCao/cLoops. TADtool is available at https://github.com/vaquerizaslab/tadtool and Juicer Tools at https://github.com/aidenlab/juicer. The other tools used are indicated in Methods. We utilized existing published analysis tools as indicated above. No new analysis tools were generated, and scripts employed to analyze the data using the published tools are available upon request.
Change history
01 November 2021
A Correction to this paper has been published: https://doi.org/10.1038/s41588-021-00980-2
References
Mayor, R. & Theveneau, E. The neural crest. Development 140, 2247–2251 (2013).
Tang, W. & Bronner, M. E. Neural crest lineage analysis: from past to future trajectory. Development 147, dev193193 (2020).
Stoller, J. Z. & Epstein, J. A. Cardiac neural crest. Semin. Cell Dev. Biol. 16, 704–715 (2005).
Vega-Lopez, G. A., Cerrizuela, S., Tribulo, C. & Aybar, M. J. Neurocristopathies: new insights 150 years after the neural crest discovery. Dev. Biol. 444, S110–S143 (2018).
Andrey, G. & Mundlos, S. The three-dimensional genome: regulating gene expression during pluripotency and development. Development 144, 3646–3658 (2017).
Rowley, M. J. & Corces, V. G. Organizational principles of 3D genome architecture. Nat. Rev. Genet. 19, 789–800 (2018).
Rao, S. S. P. et al. A 3D map of the human genome at kilobase resolution reveals principles of chromatin looping. Cell 159, 1665–1680 (2014).
Rao, S. S. P. et al. Cohesin loss eliminates all loop domains. Cell 171, 305–320.e24 (2017).
Wutz, G. et al. Topologically associating domains and chromatin loops depend on cohesin and are regulated by CTCF, WAPL, and PDS5 proteins. EMBO J. 36, 3573–3599 (2017).
Kline, A. D. et al. Diagnosis and management of Cornelia de Lange syndrome: first international consensus statement. Nat. Rev. Genet. 19, 649–666 (2018).
Piché, J., Vliet, P. P. V., Pucéat, M. & Andelfinger, G. The expanding phenotypes of cohesinopathies: one ring to rule them all! Cell Cycle 18, 2828–2848 (2019).
Chatfield, K. C. et al. Congenital heart disease in Cornelia de Lange syndrome: phenotype and genotype analysis. Am. J. Med. Genet. A 158A, 2499–2505 (2012).
Ciosk, R. et al. Cohesin’s binding to chromosomes depends on a separate complex consisting of Scc2 and Scc4 proteins. Mol. Cell 5, 243–254 (2000).
Tonkin, E. T., Wang, T.-J., Lisgo, S., Bamshad, M. J. & Strachan, T. NIPBL, encoding a homolog of fungal Scc2-type sister chromatid cohesion proteins and fly Nipped-B, is mutated in Cornelia de Lange syndrome. Nat. Genet. 36, 636–641 (2004).
Davidson, I. F. et al. DNA loop extrusion by human cohesin. Science 366, 1338–1345 (2019).
Kim, Y., Shi, Z., Zhang, H., Finkelstein, I. J. & Yu, H. Human cohesin compacts DNA by loop extrusion. Science 366, 1345–1349 (2019).
Schwarzer, W. et al. Two independent modes of chromatin organization revealed by cohesin removal. Nature 551, 51–56 (2017).
Smith, T. G. et al. Neural crest cell-specific inactivation of Nipbl or Mau2 during mouse development results in a late onset of craniofacial defects. genesis 52, 687–694 (2014).
Alesi, V. et al. Confirmation of BRD4 haploinsufficiency role in Cornelia de Lange-like phenotype and delineation of a 19p13.12p13.11 gene contiguous syndrome. Ann. Hum. Genet. 83, 100–109 (2019).
Olley, G. et al. BRD4 interacts with NIPBL and BRD4 is mutated in a Cornelia de Lange-like syndrome. Nat. Genet. 50, 329–332 (2018).
Filippakopoulos, P. et al. Selective inhibition of BET bromodomains. Nature 468, 1067–1073 (2010).
Luna-Peláez, N. et al. The Cornelia de Lange syndrome-associated factor NIPBL interacts with BRD4 ET domain for transcription control of a common set of genes. Cell Death Dis. 10, 548 (2019).
Devaiah, B. N., Gegonne, A. & Singer, D. S. Bromodomain 4: a cellular Swiss army knife. J. Leukoc. Biol. 100, 679–686 (2016).
Beliveau, B. J. et al. Versatile design and synthesis platform for visualizing genomes with Oligopaint FISH probes. Proc. Natl Acad. Sci. USA 109, 21301–21306 (2012).
Haarhuis, J. H. I. et al. The cohesin release factor WAPL restricts chromatin loop extension. Cell 169, 693–707.e14 (2017).
Tedeschi, A. et al. Wapl is an essential regulator of chromatin structure and chromosome segregation. Nature 501, 564–568 (2013).
Liu, N. Q. et al. WAPL maintains a cohesin loading cycle to preserve cell-type-specific distal gene regulation. Nat. Genet. 53, 100–109 (2021).
Kueng, S. et al. Wapl controls the dynamic association of cohesin with chromatin. Cell 127, 955–967 (2006).
Padmanabhan, A. et al. BRD4 (bromodomain-containing protein 4) interacts with GATA4 (GATA binding protein 4) to govern mitochondrial homeostasis in adult cardiomyocytes. Circulation 142, 2338–2355 (2020).
Lee, J.-E. et al. Brd4 binds to active enhancers to control cell identity gene induction in adipogenesis and myogenesis. Nat. Commun. 8, 2217 (2017).
Jiang, X., Rowitch, D. H., Soriano, P., McMahon, A. P. & Sucov, H. M. Fate of the mammalian cardiac neural crest. Development 127, 1607–1616 (2000).
Santos, R. et al. Conditional creation and rescue of Nipbl-deficiency in mice reveals multiple determinants of risk for congenital heart defects. PLoS Biol. 14, e2000197 (2016).
Ishii, M. et al. A stable cranial neural crest cell line from mouse. Stem Cells Dev. 21, 3069–3080 (2012).
Zengerle, M., Chan, K.-H. & Ciulli, A. Selective small molecule induced degradation of the BET bromodomain protein BRD4. ACS Chem. Biol. 10, 1770–1777 (2015).
Watrin, E. et al. Human Scc4 Is required for cohesin binding to chromatin, sister-chromatid cohesion, and mitotic progression. Curr. Biol. 16, 863–874 (2006).
Zhang, Q. et al. Structural mechanism of transcriptional regulator NSD3 recognition by the ET domain of BRD4. Structure 24, 1201–1208 (2016).
Shen, C. et al. NSD3-Short is an adaptor protein that couples BRD4 to the CHD8 chromatin remodeler. Mol. Cell 60, 847–859 (2015).
Jung, M. et al. Affinity map of bromodomain protein 4 (BRD4) Interactions with the histone H4 Ttail and the small molecule inhibitor JQ1*. J. Biol. Chem. 289, 9304–9319 (2014).
Crowe, B. L. et al. Structure of the Brd4 ET domain bound to a C-terminal motif from γ-retroviral integrases reveals a conserved mechanism of interaction. Proc. Natl Acad. Sci. USA 113, 2086–2091 (2016).
Konuma, T. et al. Structural mechanism of the oxygenase JMJD6 recognition by the extraterminal (ET) domain of BRD4. Sci. Rep. 7, 16272 (2017).
Hnisz, D. et al. Super-enhancers in the control of cell identity and disease. Cell 155, 934–947 (2013).
Kagey, M. H. et al. Mediator and cohesin connect gene expression and chromatin architecture. Nature 467, 430–435 (2010).
Nabet, B. et al. The dTAG system for immediate and target-specific protein degradation. Nat. Chem. Biol. 14, 431–441 (2018).
Fernandez-Alonso, R. et al. Brd4–Brd2 isoform switching coordinates pluripotent exit and Smad2-dependent lineage specification. EMBO Rep. 18, 1108–1122 (2017).
White, J. & Dalton, S. Cell cycle control of embryonic stem cells. Stem Cell Rev. 1, 131–138 (2005).
Skene, P. J. & Henikoff, S. An efficient targeted nuclease strategy for high-resolution mapping of DNA binding sites. eLife 6, e21856 (2017).
Nabet, B. et al. Rapid and direct control of target protein levels with VHL-recruiting dTAG molecules. Nat. Commun. 11, 4687 (2020).
Creyghton, M. P. et al. Histone H3K27ac separates active from poised enhancers and predicts developmental state. Proc. Natl Acad. Sci. USA 107, 21931–21936 (2010).
Lieberman-Aiden, E. et al. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science 326, 289–293 (2009).
Servant, N. et al. HiC-Pro: an optimized and flexible pipeline for Hi-C data processing. Genome Biol. 16, 259 (2015).
Dixon, J. R. et al. Topological domains in mammalian genomes identified by analysis of chromatin interactions. Nature 485, 376–380 (2012).
Nora, E. P. et al. Spatial partitioning of the regulatory landscape of the X-inactivation centre. Nature 485, 381–385 (2012).
Bonev, B. et al. Multiscale 3D genome rewiring during mouse neural development. Cell 171, 557–572.e24 (2017).
Cao, Y. et al. Accurate loop calling for 3D genomic data with cLoops. Bioinformatics 36, 666–675 (2020).
Mumbach, M. R. et al. HiChIP: efficient and sensitive analysis of protein-directed genome architecture. Nat. Methods 13, 919–922 (2016).
Zhang, H. et al. Chromatin structure dynamics during the mitosis-to-G1 phase transition. Nature 576, 158–162 (2019).
Gong, Y. et al. Stratification of TAD boundaries reveals preferential insulation of super-enhancers by strong boundaries.Nat. Commun. 9, 542 (2018).
Luppino, J. M. et al. Cohesin promotes stochastic domain intermingling to ensure proper regulation of boundary-proximal genes. Nat. Genet. 52, 840–848 (2020).
Fudenberg, G. et al. Formation of chromosomal domains by loop extrusion. Cell Rep. 15, 2038–2049 (2016).
Sanborn, A. L. et al. Chromatin extrusion explains key features of loop and domain formation in wild-type and engineered genomes. Proc. Natl Acad. Sci. USA 112, E6456–E6465 (2015).
Vian, L. et al. The energetics and physiological impact of cohesin extrusion. Cell 173, 1165–1178.e20 (2018).
Yoon, S. & Vahedi, G. Stripenn detects architectural stripes from chromatin conformation data using computer vision. Preprint at bioRxiv https://doi.org/2021.04.16.440239 (2021).
Herzog, V. A. et al. Thiol-linked alkylation of RNA to assess expression dynamics. Nat. Methods 14, 1198–1204 (2017).
Hug, C. B., Grimaldi, A. G., Kruse, K. & Vaquerizas, J. M. Chromatin architecture emerges during zygotic genome activation independent of transcription. Cell 169, 216–228.e19 (2017).
Hsieh, T.-H. S. et al. Resolving the 3D landscape of transcription-linked mammalian chromatin folding. Mol. Cell 78, 539–553.e8 (2020).
High, F. A. & Epstein, J. A. The multifaceted role of Notch in cardiac development and disease. Nat. Rev. Genet. 9, 49–61 (2008).
High, F. A. et al. An essential role for Notch in neural crest during cardiovascular development and smooth muscle differentiation. J. Clin. Invest. 117, 353–363 (2007).
Muzumdar, M. D., Tasic, B., Miyamichi, K., Li, L. & Luo, L. A global double-fluorescent Cre reporter mouse. genesis 45, 593–605 (2007).
Aghajanian, H. et al. Pdgfrα functions in endothelial-derived cells to regulate neural crest cells and the development of the great arteries. Dis. Model Mech. 10, 1101–1108 (2017).
Skene, P. J., Henikoff, J. G. & Henikoff, S. Targeted in situ genome-wide profiling with high efficiency for low cell numbers. Nat. Protoc. 13, 1006–1019 (2018).
Jones, D. T. Protein secondary structure prediction based on position-specific scoring matrices. J. Mol. Biol. 292, 195–202 (1999).
Acknowledgements
We thank the Jain and Joyce laboratories for critical discussion. We thank M. Capelson, J. A. Epstein, D. Gilbert, M. Lakadamyali, K. Musunuru and E. Nora for critical reading of the manuscript, J. E. Phillips-Cremins and M. Mir for advice, A. Silva for help with illustrations, K. Ozato for sharing the Brd4 floxed mice, A. Ciulli for sharing the MZ3 compound, A. Tsirigos and R. Sitharaman at the NYU Technology Center for Hi-C library preparation, sequencing and early informatic advice, J. Rhoades for early informatic help, B. Nabet and N. Gray for providing the dTAG compounds, D. Janssens and S. Henikoff for CUT&RUN reagents and advice, C. Petucci for metabolomic RNA labeling help, and T. Finkes, J. Haglmueller and A. Tuerk at Lexogen for SLAM-seq and analysis. This work was supported by the National Institutes of Health (NIH) (grant nos. R01 HL139783 to R.J., F31 HL147416 to R.L.-S., T32 HL007843 to B.A.K.-B., T32 GM008216 to A.K., R35 GM128903 to E.F.J., F30 HD104360 to D.S.P., T32 GM008216 to J.M.L., F31 HD102084 to J.M.L., U01 DA052715 to G.V., and K08 HL157700 to A. Padmanabhan); support from grant no. R35 HL140018 funded A. Poleshko and C.L.S.; the following also provided support: Burroughs Wellcome Foundation Career Award for Medical Scientists (R.J.), Allen Foundation (R.J.) and American Heart Association (W.K. and R.J.), the National Science Foundation (15-48571 to R.L.-S. and R.J.), Michael Antonov Charitable Foundation (A. Padmanabhan), Tobacco-Related Disease Research Foundation (A. Padmanabhan), A.P. Giannini Foundation (A. Padmanabhan) and Sarnoff Foundation (A. Padmanabhan). G.V., G.A.B., E.F.J., R.J. and their laboratories are members of the NIH 4DN consortium and thank members for critical input.
Author information
Authors and Affiliations
Contributions
R.L.-S., W.K., N.A.B. and R.J. conceived and designed the study. R.L.-S., W.K., N.A.B., B.A.K.-B., A.K. and G.H.E. performed all the experiments except for generation of the dTAG-BRD4 mESC line (Q.W. and A. Padmanabhan) and immunohistochemistry of the mouse embryo (L.L.). R.L.-S., W.K., N.A.B., H.Z., S.Y., G.H.E. and R.J. analyzed the data. R.L.-S., W.K., N.A.B., P.P.S., G.H.E. and R.J. wrote the manuscript. D.S.P., J.M.L., S.C.N., C.L.S., A. Poleshko, D.S., G.V., G.A.B. and E.F.J. assisted with data interpretation. R.J. supervised the project. All authors edited and approved the manuscript.
Corresponding author
Ethics declarations
Competing interests
D.S. is a scientific cofounder, shareholder and director of Tenaya Therapeutics. The remaining authors declare no competing interests.
Additional information
Peer review information Nature Genetics thanks the anonymous reviewers for their contribution to the peer review of this work.
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data
Extended Data Fig. 1 Characterization of BRD4 in neural crest and the interaction with NIPBL.
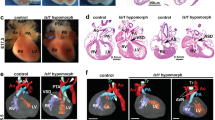
(a) GFP epifluorescence and immunohistochemistry (coronal section) of E11.5 Brd4f/f, Wnt1Cre+, R26mt-mg/+ and Brd4f/+, Wnt1Cre+, R26mt-mg/+ embryos. Scale bar, epifluorescence: 500 mm; immunohistochemistry: 500 µm. (b) Multiple sections of E16.5 Brd4f/f, Wnt1Cre+ and Brd4f/+, Wnt1Cre+ embryo showing outflow tract and heart. Mutant demonstrates right-sided aortic arch and truncus arteriosus. (Ao, Aorta; RV, right ventricle; LV, left ventricle; TA, truncus arteriosus). Second image duplicated from Fig. 1. (* denotes sectioning artifact). Scale bar, 1 mm. (c) Percentage of phospho-Histone H3 + cells in neural crest-derived craniofacial mesenchyme (each point represents one field of view, 35 (Brd4f/+, Wnt1Cre+, R26mt-mg/+) and 37 (Brd4f/f, Wnt1Cre+, R26mt-mg/+) fields of view total from two embryos, mean and SD shown, student’s T-test). (d) Expression of indicated genes in E15.0 secondary palate tissue (Unpaired T-test; ****p < 0.0001. Error bars denote SEM). Gene expression normalized to the expression of Hprt1. (e) Immunoprecipitation of endogenous BRD4 blotted for BRD4 and NIPBL (murine neural crest cells, O9-1 cells). (f) Immunoprecipitation (30 min) of endogenous BRD4 from and blotted for BRD4 and NIPBL (HEK293T cells). (g) Immunoprecipitation of endogenous BRD4 and blotted for indicated proteins (HEK293T cells). (h) Immunoprecipitation of endogenous BRD4 and blotted for indicated proteins (mESCs). (i) Indicated FLAG-tagged versions of BRD4 immunoprecipitated (HEK293T cells). Immunoblots probed with indicated antibodies. The NIPBL and FLAG immunoblots are reproduced from Fig. 2e as the proteins were all blotted for from the same blot. (j) Alignment of BRD4-interacting peptides of NSD3, LANA, JMJD6 and MLV-γ-integrase and putative BRD4-interacting peptide of NIPBL. Residues highlighted by gray are conserved across other peptides. Consensus motif indicated at the bottom (Φ represents a hydrophobic amino acid and X any amino acid). Predicted α-helix, β-strand and coil structures are marked by pink, yellow and gray color, respectively. Predictions based on PSPIRED71. The consensus motif is highlighted in the gray box.
Extended Data Fig. 2 Generation of dTAG-BRD4 mouse embryonic stem cell line.
(a) Venn diagram showing the comparison of BRD4 and NIPBL peaks from publicly available datasets (see Methods). (b) PCR of genomic DNA from clones of indicated genotype using oligos annealing outside of area of homologous recombination. “Unsuccessfully targeted” is a line which was targeted by sgRNA and Cas9, but dTAG was not inserted. Of clones shown, only Brd4dtag/dtag was confirmed by Sanger sequencing. (Ladder: Thermo, #10787-018). (c) Immunoblot of chromatin fraction from parental and dTAG-BRD4 mESCs. (d) Confirmation of fractionation protocol. Cytosol, membrane, nucleoplasm, and chromatin fractions from dTAG-BRD4 mESCs in control and the degron conditions were blotted for indicated proteins. (e) OCT4 immunofluorescence from dTAG-BRD4 mESCs in control and the degron conditions. Scale bar, 20 µm. (f) Flow cytometry-based analysis of proliferation/doubling time using CellTrace CFSE at 4.5 h post treatment with DMSO or 500 nM dTAG-13. Rectangle and circle in left and middle graphs indicates gate in which cells analyzed using Cell Proliferation tool on ModFIT LT v5.0 (n = 23823 and 23859 cells in DMSO and degron, respectively).
Extended Data Fig. 3 BRD4 regulates NIPBL occupancy.
(a) Co-occupancy analysis of BRD4 (green), NIPBL (blue) and H3K27ac (purple) at BRD4, NIPBL, or BRD4/NIPBL peaks as defined in Extended Data Fig. 2a. (b) NIPBL occupancy [log2 normalized counts (Degron/DMSO)] across top 10 K peaks. (c) Immunoblot of chromatin fractionations from DMSO, dTAG-13 (500 nM), dTAGv-1 (500 nM), and MZ3 (1 µM)-treated cells for 4.5 h for indicated proteins. The samples were derived from the same experiment and blots were processed in parallel. (d) Expression of indicated BRD4-mutants in dTAG-BRD4 mESCs upon degradation of endogenous BRD4. HDAC2 is used as a loading control. Relative band intensity ratio (BRD4:HDAC2) is indicated. (e) ChIP-qPCR of H3K27ac at regions indicated in Fig. 3e. Two-tailed T-test was used for comparison. Error bars denote the mean + SEM. (f) ChIP-qPCR of BRD4 or IgG at the same two regions assayed in Fig. 3f. One-way ANOVA Dunnett’s test was used to compare conditions to empty vector transfection with DMSO treatment. *p < 0.05. Error bars denote the mean + SD.
Extended Data Fig. 4 BRD4 maintains higher order chromatin structure.
(a) Hi-C matrix comparing c-loop identified loops in DMSO, dTAG-BRD4 mESCs (blue) to mESCs profiled in Mumbach et al., 2016 (black) at two different regions. (b) Mean counts/bin of gained interactions in DMSO and the degron conditions. T-test used for comparison. (c) NIPBL occupancy at BRD4-sensitive and -resistant loop anchors in DMSO and the degron conditions. (d) Stratification of BRD4-resistant and -sensitive loops. Structural loops refer to loops with CTCF and RAD21 peaks at both anchors and 0-1 anchors have H3K27ac peak. CTCF-Cohesin enhancer-promoter (EP) loops refer to loops with H3K27ac, CTCF and RAD21 peaks at both anchors. CTCF-Cohesin independent loops refer to loops with CTCF and RAD21 peaks at 0-1 anchors and both anchors have H3K27ac peak. Other refers to all loops which do not fall into these categories. Anchor regions were extended by 15 kb upstream and downstream for peak intersections. (e) Change in interaction frequency at BRD4, NIPBL, RAD21, and CTCF peaks. (f) Change in interaction frequency at NIPBL peaks (stratified into deciles of peak strength) upon depletion of BRD4 (left) and BRD4 occupancy (normalized counts) at NIPBL peaks (right) (stratified into deciles of peak strength). (g) Hi-C matrix comparing TADs in DMSO, dTAG-BRD4 mESCs (blue) to mESCs profiled in Dixon et al.51 (black) at two different regions. Adjacent bar plot compares the percentage of basepairs uniquely found in a TAD in DMSO, dTAG-BRD4 mESCs (light blue), Dixon et al.51 mESCs (black) or shared between the datasets (navy). Jaccard index between datasets 0.82. (h) Correlation of insulation score specifically at TAD boundaries between DMSO and the degron conditions.
Extended Data Fig. 5 BRD4 loss does not affect compartmentalization.
(a) Eigenvalues in 250 kb bins across the entire genome in DMSO (blue) and the degron (red) conditions, per chromosome. (b) Genome wide comparison of eigenvector values (250 kb bins) in DMSO (blue) and the degron (red) conditions. (c) Correlation of eigenvector values between DMSO and the degron conditions. T-test used for comparison.
Extended Data Fig. 6 Loss of BRD4 abrogates genome folding.
(a) Hi-C matrix (10 kb resolution) of region painted with Oligopaints (as indicated by the green and magenta bars below matrix, from Fig. 5a). Below matrix is insulation score and NIPBL occupancy in DMSO and degron conditions. (b) Immunoblot of NIPBL in shScramble and shNipbl treatment in mESCs. HDAC2 was blotted as a loading control. (c) FISH signal compactness (see methods) of upstream and downstream probes used in Fig. 5. (d) Hi-C matrix (10 kb resolution) with areas painted with Oligopaints, insulation score, BRD4 and NIPBL occupancy (in wildtype mESCs), NIPBL occupancy in DMSO and degron samples shown below. Area highlighted by red dotted line indicates sub-TAD and arrows point to a domain affected upon BRD4 depletion. On right is a representative image from DMSO and the degron conditions with regions painted indicated below matrix. Below the representative FISH images is the plot of center-to-center signal distances and compactness of FISH signals. For visualization purposes only, only probe center-to-center distances up to 0.6 μm shown. Wilcoxon-rank sum test. Scale bar, 1 µm. (e) Hi-C matrix (10 kb resolution) with areas painted with Oligopaints, insulation score, BRD4 and NIPBL occupancy (in wildtype mESCs), and NIPBL occupancy in DMSO and degron samples shown below. In middle is representative image from DMSO and the degron conditions with regions indicated on matrix painted. On right is the plot of center-to-center distances. Wilcoxon-rank sum test used for comparison. Scale bar, 1 µm. All images are maximum-projections. (f) Representative GFP immunofluorescence from conditions used in Fig. 5c. Scale bar, 100 µm. (g) ChIP-qPCR of BRD4 or IgG at the two regions indicated in Fig. 5a by red bars. One-way ANOVA Dunnett’s test was used to compare conditions to empty vector transfection with DMSO treatment. *p < 0.05. Error bars denote the mean + SD.
Extended Data Fig. 7 BRD4 mediates loop extrusion.
(a) Chromatin fractionation immunoblot of dTAG-BRD4 mESCs treated with DMSO, dTAG-13 (500 nM), dTAGv-1 (500 nM), or MZ3 (1 µM) for 4.5 h. The NIPBL, BRD4 and HDAC2 immunoblots are reproduced from Extended Data Fig. 2f as the proteins were all blotted from the same blot. The samples were derived from the same experiment and blots were processed in parallel. (b) Occupancy from public datasets (see methods) of indicated factors (black) and random shuffle (gray) across stripe domains in DMSO-treated dTAG-BRD4 mESCs. Note the enrichment of factor occupancy at stripe anchors. (c) Hi-C matrix (top DMSO, bottom Degron) showing an example of a stripe domains (10 kb resolution), which is magnified on the right. Black arrows indicate a stripe domain which is weaker in degron compared to control. Color bar refers normalized count. (d) Hi-C matrix (top DMSO, bottom Degron) showing another example of stripe (indicated by arrow) which is weaker in degron compared to DMSO (10 kb resolution). (e) Change in stripe score and median intensity of stripe domains in degron compared to DMSO. Stripe domains defined in DMSO condition and same region compared in the degron condition. T-test used for comparison. (f–i) Occupancy of NIPBL (F and G) or RAD21 (H and I) at RAD21-NIPBL peaks (F and H) or in stripe domains (G-NIPBL peaks and I-RAD21 peaks) in DMSO and the degron conditions. (j) The percentage of differentially expressed genes per TAD is plotted and TADs are broken into descending deciles of change in interaction frequency. Individual data points shown as well as superimposed box and whisker plot (median and interquartile range indicated by box). (k) Cumulative density of differentially expressed genes (DEG, FDR < 0.05, |log2FC | >1, red) or all genes (black) in relation to proximity to BRD4-sensitive (left) and -resistant (right) loop anchors.
Extended Data Fig. 8 BRD4 mediates differentiation of neural crest progenitor cells.
(a) Immunoblot showing levels of BRD4 upon depletion of BRD4 and introduction of wildtype BRD4 or various mutants unable to bind NIPBL. EV: empty vector. Undif.: Undifferentiated condition. (b) Representative image of undifferentiated O9-1 cells infected with shRenilla and stained with SMA antibody. Scale bar, 100 μm. (c) Acta2 and Cnn1 expression in indicated conditions. One-way ANOVA Tukey’s multiple comparison test is used to compare across conditions (*p < 0.05, ***p < 0.001). Error bars denote the mean ± SD. Undif.: Undifferentiated condition. (d) Nipbl expression in Brd4-depleted O9-1 cells. Unpaired t-test was used for comparison. All error bars denote the mean ± SEM. (e) Immunoblots of BRD4, WAPL and NIPBL in various knockdown conditions. Histone H3 and GAPDH are loading controls. Undif.: Undifferentiated condition. (f) Representative image of undifferentiated O9-1 cells infected with shRenilla and transfected with empty vector and stained with SMA antibody. Scale bar, 100 μm. (g, h) Acta2 and Cnn1 expression in indicated conditions. One-way ANOVA Tukey’s multiple comparison test is used to compare across conditions (*p < 0.05, **p < 0.01, ***p < 0.001). Error bars denote the mean ± SD. Undif.: Undifferentiated condition.
Supplementary information
Supplementary Information
Supplementary discussion, methods and references.
Supplementary Tables
Supplementary Tables 1–6.
Source data
Source Data Fig. 2
Unprocessed western blots and/or gels.
Source Data Fig. 3
Unprocessed western blots and/or gels.
Source Data Extended Data Fig. 1
Unprocessed western blots and/or gels.
Source Data Extended Data Fig. 2
Unprocessed western blots and/or gels.
Source Data Extended Data Fig. 3
Unprocessed western blots and/or gels.
Source Data Extended Data Fig. 6
Unprocessed western blots and/or gels.
Source Data Extended Data Fig. 7
Unprocessed western blots and/or gels.
Source Data Extended Data Fig. 8
Unprocessed western blots and/or gels.
Rights and permissions
About this article

Cite this article
Linares-Saldana, R., Kim, W., Bolar, N.A. et al. BRD4 orchestrates genome folding to promote neural crest differentiation. Nat Genet 53, 1480–1492 (2021). https://doi.org/10.1038/s41588-021-00934-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41588-021-00934-8
This article is cited by
-
Taking BETs on cardiomyocyte differentiation
Nature Cardiovascular Research (2024)
-
A genome-wide CRISPR screen identifies BRD4 as a regulator of cardiomyocyte differentiation
Nature Cardiovascular Research (2024)
-
Chromatin remodeler Activity-Dependent Neuroprotective Protein (ADNP) contributes to syndromic autism
Clinical Epigenetics (2023)
-
High-sensitive nascent transcript sequencing reveals BRD4-specific control of widespread enhancer and target gene transcription
Nature Communications (2023)
-
The Mediator complex regulates enhancer-promoter interactions
Nature Structural & Molecular Biology (2023)
