
Abstract
The shaping of human embryos begins with compaction, during which cells come into close contact1,2. Assisted reproductive technology studies indicate that human embryos fail compaction primarily because of defective adhesion3,4. On the basis of our current understanding of animal morphogenesis5,6, other morphogenetic engines, such as cell contractility, could be involved in shaping human embryos. However, the molecular, cellular and physical mechanisms driving human embryo morphogenesis remain uncharacterized. Using micropipette aspiration on human embryos donated to research, we have mapped cell surface tensions during compaction. This shows a fourfold increase of tension at the cell–medium interface whereas cell–cell contacts keep a steady tension. Therefore, increased tension at the cell–medium interface drives human embryo compaction, which is qualitatively similar to compaction in mouse embryos7. Further comparison between human and mouse shows qualitatively similar but quantitively different mechanical strategies, with human embryos being mechanically least efficient. Inhibition of cell contractility and cell–cell adhesion in human embryos shows that, whereas both cellular processes are required for compaction, only contractility controls the surface tensions responsible for compaction. Cell contractility and cell–cell adhesion exhibit distinct mechanical signatures when faulty. Analysing the mechanical signature of naturally failing embryos, we find evidence that non-compacting or partially compacting embryos containing excluded cells have defective contractility. Together, our study shows that an evolutionarily conserved increase in cell contractility is required to generate the forces driving the first morphogenetic movement shaping the human body.
This is a preview of subscription content, access via your institution
Access options
Access Nature and 54 other Nature Portfolio journals
Get Nature+, our best-value online-access subscription
$29.99 / 30 days
cancel any time
Subscribe to this journal
Receive 51 print issues and online access
$199.00 per year
only $3.90 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout



Similar content being viewed by others


Data availability
Images and data are available from the BioImage Archive under accession number S-BIAD915. Simulations of compaction using surface tension values from experiments were used to illustrate different scenarios of surface tension changes. Source data are provided with this paper.
Code availability
Custom code is available at https://doi.org/10.5281/zenodo.10779533 (ref. 45).
References
Firmin, J. & Maître, J.-L. Morphogenesis of the human preimplantation embryo: bringing mechanics to the clinics. Semin. Cell Dev. Biol. 120, 22–31 (2021).
Shahbazi, M. N. Mechanisms of human embryo development: from cell fate to tissue shape and back. Development 147, dev190629 (2020).
Coticchio, G., Lagalla, C., Sturmey, R., Pennetta, F. & Borini, A. The enigmatic morula: mechanisms of development, cell fate determination, self-correction and implications for ART. Hum. Reprod. Update 25, 422–438 (2019).
Lagalla, C. et al. Embryos with morphokinetic abnormalities may develop into euploid blastocysts. Reprod. BioMed. Online 34, 137–146 (2017).
Collinet, C. & Lecuit, T. Programmed and self-organized flow of information during morphogenesis. Nat. Rev. Mol. Cell Biol. 22, 245–265 (2021).
Heisenberg, C.-P. & Bellaïche, Y. Forces in tissue morphogenesis and patterning. Cell 153, 948–962 (2013).
Maître, J.-L., Niwayama, R., Turlier, H., Nédélec, F. & Hiiragi, T. Pulsatile cell-autonomous contractility drives compaction in the mouse embryo. Nat. Cell Biol. 17, 849–855 (2015).
Haniffa, M. et al. A roadmap for the Human Developmental Cell Atlas. Nature 597, 196–205 (2021).
Rossant, J. & Tam, P. P. L. Opportunities and challenges with stem cell-based embryo models. Stem Cell Rep. 16, 1031–1038 (2021).
Özgüç, Ö. & Maître, J.-L. Multiscale morphogenesis of the mouse blastocyst by actomyosin contractility. Curr. Opin. Cell Biol. 66, 123–129 (2020).
Fogarty, N. M. E. et al. Genome editing reveals a role for OCT4 in human embryogenesis. Nature 550, 67–73 (2017).
Gerri, C. et al. Initiation of a conserved trophectoderm program in human, cow and mouse embryos. Nature 587, 443–447 (2020).
Okamoto, I. et al. Eutherian mammals use diverse strategies to initiate X-chromosome inactivation during development. Nature 472, 370–374 (2011).
Petropoulos, S. et al. Single-cell RNA-seq reveals lineage and X chromosome dynamics in human preimplantation embryos. Cell 165, 1012–1026 (2016).
Iwata, K. et al. Analysis of compaction initiation in human embryos by using time-lapse cinematography. J. Assist. Reprod. Genet. 31, 421–426 (2014).
Coticchio, G. et al. Perturbations of morphogenesis at the compaction stage affect blastocyst implantation and live birth rates. Hum. Reprod. 36, 918–928 (2021).
Rienzi, L. et al. Time of morulation and trophectoderm quality are predictors of a live birth after euploid blastocyst transfer: a multicenter study. Fertil. Steril. 112, 1080–1093 (2019).
Skiadas, C., Jackson, K. & Racowsky, C. Early compaction on day 3 may be associated with increased implantation potential. Fertil. Steril. 86, 1386–1391 (2006).
Turlier, H. & Maître, J.-L. Mechanics of tissue compaction. Semin. Cell Dev. Biol. 47–48, 110–117 (2015).
Goel, N. S., Doggenweiler, C. F. & Thompson, R. L. Simulation of cellular compaction and internalization in mammalian embryo development as driven by minimization of surface energy. Bull. Math. Biol. 48, 167–187 (1986).
Pelzer, D. et al. Cell fragmentation in mouse preimplantation embryos induced by ectopic activation of the polar body extrusion pathway. EMBO J. 42, e114415 (2023).
Chugh, P. et al. Actin cortex architecture regulates cell surface tension. Nat. Cell Biol. 19, 689–697 (2017).
Özgüç, Ö. et al. Cortical softening elicits zygotic contractility during mouse preimplantation development. PLoS Biol. 20, e3001593 (2022).
Schliffka, M. F. et al. Multiscale analysis of single and double maternal-zygotic Myh9 and Myh10 mutants during mouse preimplantation development. eLife 10, e68536 (2021).
Maître, J.-L. & Heisenberg, C.-P. Three functions of cadherins in cell adhesion. Curr. Biol. 23, R626–R633 (2013).
Yan, L. et al. Single-cell RNA-seq profiling of human preimplantation embryos and embryonic stem cells. Nat. Struct. Mol. Biol. 20, 1131–1139 (2013).
Wamaitha, S. E. & Niakan, K. K. Human pre-gastrulation development. Curr. Top. Dev. Biol. 128, 295–338 (2018).
Zakharova, E. E., Zaletova, V. V. & Krivokharchenko, A. S. Biopsy of human morula-stage embryos: outcome of 215 IVF/ICSI cycles with PGS. PLoS ONE 9, e106433 (2014).
Maitre, J.-L. et al. Adhesion functions in cell sorting by mechanically coupling the cortices of adhering cells. Science 338, 253–256 (2012).
Chan, E. H., Chavadimane Shivakumar, P., Clément, R., Laugier, E. & Lenne, P.-F. Patterned cortical tension mediated by N-cadherin controls cell geometric order in the Drosophila eye. eLife 6, e22796 (2017).
Stirbat, T. V. et al. Fine tuning of tissues’ viscosity and surface tension through contractility suggests a new role for α-catenin. PLoS ONE 8, e52554 (2013).
Guck, J. Some thoughts on the future of cell mechanics. Biophys. Rev. 11, 667–670 (2019).
Budczies, J. et al. Cutoff Finder: a comprehensive and straightforward web application enabling rapid biomarker cutoff optimization. PLoS ONE 7, e51862 (2012).
Coorens, T. H. H. et al. Inherent mosaicism and extensive mutation of human placentas. Nature 592, 80–85 (2021).
Maître, J.-L. et al. Asymmetric division of contractile domains couples cell positioning and fate specification. Nature 536, 344–348 (2016).
Matamoro-Vidal, A. & Levayer, R. Multiple influences of mechanical forces on cell competition. Curr. Biol. 29, R762–R774 (2019).
True, J. R. & Haag, E. S. Developmental system drift and flexibility in evolutionary trajectories. Evol. Dev. 3, 109–119 (2001).
Ichbiah, S., Delbary, F., McDougall, A. & Dumollard, R. Embryo mechanics cartography: inference of 3D force atlases from fluorescence microscopy. Nat. Methods 20, 1989–1999 (2023).
Lenne, P.-F. et al. Roadmap for the multiscale coupling of biochemical and mechanical signals during development. Phys. Biol. 18, 041501 (2021).
Clark, A. T. et al. Human embryo research, stem cell-derived embryo models and in vitro gametogenesis: considerations leading to the revised ISSCR guidelines. Stem Cell Rep. 16, 1416–1424 (2021).
Tsunoda, Y., Yasui, T., Nakamura, K., Uchida, T. & Sugie, T. Effect of cutting the zona pellucida on the pronuclear transplantation in the mouse. J. Exp. Zool. 240, 119–125 (1986).
Guevorkian, K. & Maître, J.-L. Micropipette aspiration: a unique tool for exploring cell and tissue mechanics in vivo. Methods Cell. Biol. 139, 187–201 (2017).
Schindelin, J. et al. Fiji: an open-source platform for biological-image analysis. Nat. Methods 9, 676–682 (2012).
Alpha Scientists in Reproductive Medicine and ESHRE Special Interest Group of Embryology et al. The Istanbul consensus workshop on embryo assessment: proceedings of an expert meeting. Hum. Reprod. 26, 1270–1283 (2011).
Ecker, N. & Turlier, H. 3D active foam simulations. Zenodo https://zenodo.org/doi/10.5281/zenodo.10779532 (2024).
Acknowledgements
We thank the imaging platform of the Genetics and Developmental Biology unit at the Institut Curie (PICT-IBiSA@BDD), member of the French National Research Infrastructure France-BioImaging (ANR-10-INBS-04) for their outstanding support. We thank N. Kazdar, L. Delaroche and A. Le Dû and all the members of ART teams from the Clinique La Muette (Paris, France), the Clinique Pierre Chérest (Neuilly sur Seine, France) and the Hopital Cochin (Paris, France) for support with human embryo experiments. We thank all members of the Maître laboratory, Y. Bellaïche and M.-H. Verlhac for discussion and comments. We acknowledge the support with administrative issues from M.-H. Verlhac throughout this project. We are grateful to the patients who donated their surplus embryos to research. This project was funded by a Paris Sciences et Lettres (PSL) QLife (ANR-17-CONV-0005) grant to J.-L.M., C.P. and H.T. and the INSERM transversal programme Human Development Cell Atlas (HuDeCA) to J.-L.M. and C.P. Research in the laboratory of J.-L.M. is supported by the Institut Curie, the Centre National de la Recherche Scientifique (CNRS), the Institut National de la Santé Et de la Recherche Médicale (INSERM) and is funded by grants from the Fondation Schlumberger pour l’Éducation et la Recherche through the Fondation pour la Recherche Médicale, the European Molecular Biology Organization Young Investigator program (EMBO YIP), Labex DEEP (ANR- 11-LABX-0044, part of the IDEX PSL ANR-10-IDEX-0001–02). J.F. is funded by a fellowship from the Fondation pour la Recherche Médicale (FDM202006011290). The work by H.T. and N.E. was supported by the CNRS and Collège de France. No fund from the European Research Council was used for this project.
Author information
Authors and Affiliations
Contributions
J.F., H.T., C.P. and J.-L.M. conceptualized the project and acquired funding. J.F. and J.-L.M. designed experiments. J.F. performed experiments. J.F., Ö.Ö. and J.-L.M. analysed the data. J.F., D.R.D., V.B.L and C.P. organized embryo collection. N.E. and H.T. wrote the theory and performed numerical simulations. J.-L.M. wrote the manuscript with inputs from J.F., N.E., H.T. and C.P.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature thanks Berenika Plusa, Hilde Van de Velde, Alpha Yap and the other, anonymous, reviewer(s) for their contribution to the peer review of this work. Peer reviewer reports are available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
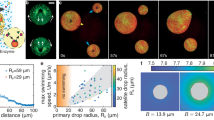
Extended Data Fig. 1 Contact angle and surface tension measurements of individual embryos.
(a-c) Time course of external contact angles θe (a) and surface tensions γcm (b) and γcc/2 (c) of 5 representative embryos. Time starts from the first tension measurement. Each embryo is measured in 3–5 sessions, for which the mean ± SEM of 2 to 12 contacting cells are shown. (d-e) Mean ± SEM surface tension γcm (d) and γcc/2 (e) as a function of contact angles θe over successive measurement session of 5 representative embryos. Each embryo is measured in 3–5 sessions, for which the mean ± SEM of 2 to 12 contacting cells are shown. (f-g) Surface tension γcm (f) and γcc/2 (g) as a function of contact angles θe measured on 429 blastomeres from 14 embryos. Individual measurements are shown in grey (Pearson correlation values R = 0.624 for γcm (p < 10−47) and R = −0.135 for γcc (p > 10−2)) and mean ± SEM of 44 individual measurement sessions on 14 embryos are shown in red for γcm (Pearson correlation values of R = 0.740 (p < 10−8)) and in green for γcc R = 0.028 for γcc (p > 10−1)). p values obtained from the 2-tailed Pearson correlation critical values table.
Extended Data Fig. 2 Cleavage stage of compacting and non-compacting blastomeres.
Blastomere stage, as determined by tracking successive cell divisions until the last tension measurement, of compacting, non-compacting and partially compacting embryos (7, 6 and 7 embryos and 40, 32 and 33 blastomeres respectively). For partially compacting embryos, the stages of compacted blastomeres, compacted blastomeres adjacent to excluded cells and excluded cells are indicated separately (13, 10 and 10 respectively).
Extended Data Fig. 3 Volume and pressure changes during compaction.
(a-b) Hydrostatic pressure (a) and radius of curvature (b) of cells as a function of contact angles θe measured on 429 blastomeres from 14 embryos. Cleavage stages are determined based on tracking divisions on time lapse microscopy and are indicated with 4-, 8- and 16-cell stage blastomeres in grey, light and dark blue respectively. Black dots show blastomeres that cannot be staged with certainty. Pearson correlation R = 0.551 for the hydrostatic pressure and −0.044 for the radius of curvature. (c) Volume segmentation of embryos before and after compaction. Cells are shown in light blue when estimated to be the size of a 8-cell stage blastomere and larger, or shown in dark blue when the size of a 16-cell stage blastomere and smaller. (d) Volume of segmented cells as a function of the total number of cells in the embryo (118 cells from 21 embryos). Embryos with precisely 8 or 16 cells are indicated in light or dark blue respectively. Embryos with precisely 8 or 16 cells are used to statistically determine the characteristic size of 8- and 16-cell stage blastomeres respectively. Using these measurements, a size threshold is statistically determined at 49850 µm3 to classify cells as 8-cell stage blastomere and larger or 16-cell stage blastomere and smaller33. (e) Volume of 118 segmented cells as a function of the contact angle θe measured on 112 contacts from 21 embryos. Cells classified as 8-cell stage blastomere and larger are shown in light blue and cells classified as 16-cell stage blastomere and smaller are shown in dark blue. Pearson correlation between cell volume and contact angles are 0.176 for 8-cell stage blastomeres and larger (59 contacts, p > 10−1) and −0.034 for 16-cell stage blastomeres and smaller (53 contacts, p > 10−1). p values obtained from 2-tailed Pearson correlation tests.
Extended Data Fig. 4 Recovery of compaction after ML7 and EDTA media treatments.
(a) Representative images of embryos in control medium (left) placed in medium containing 1:2000 ethanol (EtOH) for 45 min, then 10 µM ML7 for 45 min and after 3 h recovery in control medium (right). Scale bar, 20 µm. (b) Contact angles θe of embryos placed sequentially in control EtOH, ML7 and control media (Mean ± SEM of 59, 51, 44 contacts from 6 embryos). (c) Representative images of embryos placed in control medium (left) for 45 min, then EDTA containing medium for 30 min (center) and after 3 h recovery in control medium (right). Scale bar, 20 µm. (d) Contact angles θe of embryos placed sequentially in control, EDTA and control media (Mean ± SEM of 54, 43, 54 contacts from 6 embryos).
Supplementary information
Supplementary Information
Supplementary table legends, video legends and references.
Supplementary Tables
Supplementary Tables 1–8.
Supplementary Video 1
Pre-implantation development of human embryos with full compaction. Time-lapse imaging of a human embryo from the four-cell stage to the blastocyst stage (shown in Fig 1b). Pictures taken every 30 min; scale bar, 40 µm.
Supplementary Video 2
Pre-implantation development of human embryos without compaction. Time-lapse imaging of a human embryo from the four-cell stage to the 16-cell stage (shown in Fig 3a). Pictures taken every 30 min; scale bar, 40 µm.
Supplementary Video 3
Pre-implantation development of human embryos with partial compaction. Time-lapse imaging of a human embryo from the four-cell stage to the blastocyst stage (shown in Fig 3e). Pictures taken every 30 min; scale bar, 40 µm.
Supplementary Video 4
Three-dimensional simulations of compaction. Simulations of compaction with distinct cell populations: blue blastomeres grow their tension γcm by a factor 3.2 and their tension γcc by 1.2 according to measurements shown in Fig. 3i,j; purple blastomeres do the same as blue ones (left), keep γcm steady and grow their tension γcc by 1.2 (middle) or keep γcm steady and grow their tension γcc by 2.4 (right). Tensions are linearly interpolated between the initial and final states in 15 steps.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Firmin, J., Ecker, N., Rivet Danon, D. et al. Mechanics of human embryo compaction. Nature 629, 646–651 (2024). https://doi.org/10.1038/s41586-024-07351-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41586-024-07351-x
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
