
Abstract
Groundwater is critical for many ecosystems, yet groundwater requirements for dependent ecosystems are rarely accounted for during water and conservation planning. Here we compile 38 years of Landsat-derived normalized difference vegetation index (NDVI) to evaluate groundwater-dependent vegetation responses to changes in depth to groundwater (DTG) across California. To maximize applicability, we standardized raw NDVI and DTG values using Z scores to identify groundwater thresholds, groundwater targets and map potential drought refugia across a diversity of biomes and local conditions. Groundwater thresholds were analysed for vegetation impacts where ZNDVI dropped below −1. ZDTG thresholds and targets were then evaluated with respect to groundwater-dependent vegetation in different condition classes and rooting depths. ZNDVI scores were applied statewide to identify potential drought refugia supported by groundwater. Our approach provides a simple and robust methodology for water and conservation practitioners to support ecosystem water needs so biodiversity and sustainable water-management goals can be achieved.
Similar content being viewed by others
Main
Groundwater is a critical component of many aquatic and terrestrial ecosystems, but its role in sustaining ecosystem function and stability is rarely acknowledged in conservation and water-management planning globally1,2,3. Increasing evidence over recent decades has linked groundwater pumping and other water-use practices to adverse impacts on groundwater-dependent species, habitats and ecosystems through declines in streamflow4,5,6 and groundwater levels7. Moreover, rates of groundwater pumping are likely to intensify as global warming increases the frequency and severity of extreme drought events8,9,10. Subsequent groundwater declines will amplify adverse ecosystem impacts because groundwater is an essential buffer in meeting higher evapotranspiration demands under drought11,12,13. Groundwater-related adverse impacts range in severity from water stress (individual scale) to habitat loss (population scale) to, in the worst-case scenario, ecosystem collapse (system scale). Whereas some impacts may be reversible, others may result in the permanent loss of species and/or habitats. To meet global biodiversity and sustainable water-management goals, while preserving cultural values associated with these ecosystems, ecosystem water requirements need to be identified and quantified1,14,15. However, ecological thresholds and targets for groundwater are not well defined due to deficiencies in data, ecohydrologic understanding, legal protections or political will in water-management agencies2,16,17,18,19. Even where targets exist, traditional approaches to adaptive resource management may not be appropriate due to high levels of uncertainty and the potential severity and permanence of impacts from groundwater-management actions20. Thus, a practical approach for quantifying ecosystem groundwater requirements across a wide range of ecosystem and local conditions is needed to support conservation action, water-management and water-allocation decisions.
Groundwater-dependent ecosystems (GDEs) are comprised of species and habitats that rely on groundwater for some or all of their water needs. These ecosystems are incredibly diverse, existing across above- and below-ground aquatic and terrestrial realms and supporting a wide range of species and niche habitats19. Whereas groundwater can support subterranean ecosystems (for example, in caves, aquifers)3, our study focuses on terrestrial groundwater-dependent ecosystems with perennial vegetation, including riparian woodlands that are reliant on groundwater occurring on or near Earth’s surface. Rather than measuring the functional biological responses of all taxonomic groups within these ecosystems, here we target the dominant groundwater-dependent vegetation (‘phreatophyte’) species and monitor their sensitivity to changes in groundwater availability. Phreatophytes are prominent members of many groundwater-dependent ecosystems and are not only good indicators of near-surface groundwater conditions7 but are also considered ‘foundation’ species within their communities21. Woody phreatophytes, such as the dominant trees in riparian ecosystems, provide structure, biomass, material flows and microclimate regulation for many dependent species. Canopy stratification and vertical complexity creates diverse habitats that support associated fauna species including birds, mammals, fish and insects. Phreatophytes can also be monitored using remote sensing methods to detect responses over large areas. Phreatophytes are thus appropriate and practical indicators for evaluating ecosystem groundwater requirements across a region, improving the applicability of ecosystem groundwater needs assessments in water-use and planning decisions.
Ecosystem groundwater needs assessments require science-based ecological thresholds that quantify the transition between functionally stable and detrimental ecosystem states and science-based ecological targets that quantify ecosystem states that have the physiological capacity to deal with natural range of hydrologic variability14. Previous studies have demonstrated that both the rate of water table decline and absolute groundwater depth can trigger declines in vegetation health12,13. This is because phreatophyte species can often tolerate or adapt to gradual changes in groundwater levels, but if these changes are prolonged or abrupt, it can result in a range of functional or ecological responses ranging in severity from declines in productivity, recruitment and mortality22,23,24,25. Thus, groundwater thresholds and targets must take into consideration the rate, magnitude and duration of the groundwater change and how these relate to either temporary or long-term impacts on an ecosystem26,27. Therefore, determining ecosystem groundwater requirements depends upon understanding groundwater-level thresholds and targets required to sustain groundwater-dependent ecosystems, so that management actions can augment supplies (for example, managed aquifer recharge projects) or reduce demand (for example, pumping restrictions) to maintain ecological function and sustain species and their communities.
The challenge is that vegetation responses to groundwater changes can vary substantially over time and space due to natural seasonal and interannual groundwater fluctuations, species-specific tolerance and adaptations to water stress and the existence of supplemental water sources such as surface water, irrigation return flow and treated wastewater effluent28. Furthermore, the effects of groundwater changes can vary even within species due to local vegetation density and biomass that influence demand and individuals’ adaptation to site-specific hydrology and climate during establishment and growth. These uncertainties are further complicated by the impacts of climate change on the water cycle and vegetation, as well as existing groundwater data gaps in most regions7,28.
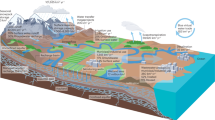
To address this challenge, we utilize Z scores as a means to standardize vegetation responses and groundwater levels for quantifying ecological thresholds and targets relative to a historic baseline that can be consistently applied across a diversity of ecosystems and local environmental conditions (Fig. 1). Z scores are a well-established statistical metric to indicate how many standard deviations an individual observation value is from the mean and are useful in comparing values from distributions with different units and ranges. Thus, Z scores calculated from the range of location-specific values enable us to track ecological responses to groundwater levels that fluctuate above and below local baseline conditions, which can vary greatly due to vegetation species composition and density, aquifer properties, natural climate and surface water flow and groundwater regimes.

a, Landscape schematic diagram illustrating how normalized difference vegetation index (NDVI) and depth-to-groundwater (DTG) fluctuations can vary temporally and spatially across the landscape depending on land surface elevation and landscape positioning. Groundwater depths and associated NDVI values are site specific and provided here for illustrative purposes. Most vegetation are adapted to natural fluctuations in DTG (for example, within −1 to +1 standard deviation (σ) over a baseline period), but if DTG exceeds these natural fluctuations, vegetation can become adversely impacted. b, DTG naturally fluctuates over time, but drought events and intensive groundwater pumping can cause groundwater levels to extend far below the natural range of variability observed over time. c, Z scores can be used to standardize groundwater levels as an alternative metric for quantifying ecological thresholds and targets across environments with variable local conditions.
Here we evaluate ecological thresholds, ecological targets and map potential drought refugia for groundwater-dependent vegetation across California (n = 246,017 vegetation polygons), where state law requires an evaluation of impacts to ecosystems under groundwater law (Sustainable Groundwater Management Act) and the Public Trust Doctrine. We base this analysis on 38 years (1985–2022) of 30-m resolution Landsat-derived normalized difference vegetation index (NDVI) data (n = 9,343,646 annual dry-season observations) to monitor vegetation responses to groundwater changes over time. To identify groundwater thresholds resulting in adverse GDE impacts, we analysed average annual dry-season (July–September) NDVI and field-based groundwater levels (n = 36,381 paired observations) in California where shallow groundwater data exist. Dry-season groundwater and NDVI data were selected because reliance on groundwater is generally higher during drier months when soil moisture from surface water and precipitation is scarce, supporting vegetation to maintain vigour later into the season28,29. We transformed both the NDVI and depth-to-groundwater (DTG) data to Z scores using the distribution of values for each polygon to compute the statistical moments, thus standardizing the response values according to the local setting. This approach evaluates acute groundwater threshold responses that are visible from satellite imagery such as reduction in greenness, severe vegetation die-back and mortality, as observed during the 2012–2015 drought12,13,30 (Fig. 2). However, chronic responses or subtle community responses to prolonged groundwater decline such as species shifts or declines in recruitment are not thoroughly explored here due to data limitations. We evaluate ecologic targets by comparing groundwater levels to rooting depths for plant genera common to groundwater-dependent ecosystems with respect to their greenness (NDVI) Z scores. Lastly, we infer potential hydrologic refugia across California where groundwater data are lacking by utilizing the >9.3 million ZNDVI scores (Supplementary Section 2) to map where vegetation greenness was stable during dry years and multi-year droughts. Finally, we discuss the implications of our findings for water resource management and drought response in California and beyond.

a,b, Drought-impacted groundwater-dependent vegetation as indicated by the change in NDVI (max–min) during the 2012–2015 drought (a) and visible from National Agriculture Imagery Program (NAIP) imagery at select locations (b): Salinas River (gde_id: 35979, outlined in orange), Santa Clara River (gde_id: 111026, outlined in orange), Kern River (gde_id: 134058, outlined in orange).
Results
Drought-impacted vegetation provide insight into thresholds
We analysed negative impacts to groundwater-dependent vegetation throughout California where associated groundwater data exist (Fig. 3). Groundwater data were available for only 1.3% (n = 3,113 polygons) of California’s groundwater-dependent vegetation. In contrast to plotting the raw NDVI and DTG values, whose mean value and range can vary greatly depending on local conditions (Fig. 4 and Supplementary Fig. 2), the Z score plots account for large differences in average or baseline conditions (for example, local vegetation density in the case of NDVI or mean water table depth for DTG) and the range of variability experienced locally and to which the vegetation is adapted. Thus, there was a stronger negative relationship between ZNDVI and ZDTG than between the raw NDVI and DTG values, indicating lower greenness values with deeper groundwater for all vegetation and individual genera (Fig. 4). The linear regression between ZNDVI and ZDTG for vegetation associated with shallow monitoring wells along the Kern River exhibited a tighter linear fit than the California Statewide Groundwater Elevation Monitoring (CASGEM) monitoring data, indicating that some uncertainty exists in the statewide groundwater data, because most CASGEM monitoring wells were not always specifically positioned to monitor shallow groundwater conditions along riparian corridors, wetlands or other groundwater-dependent ecosystems (Methods). The ZNDVI and ZDTG relationship for three major groundwater-dependent vegetation genera across California (Salix, Populus and Quercus) also exhibited significant negative correlations with Quercus being the least sensitive in response to water table changes. This is probably due to Quercus trees having a deeper rooting structure enabling it to access deeper groundwater31 and water-stress-coping mechanisms such as hydraulic redistribution and water efficiency resulting in greater plasticity in response to groundwater decline32,33 relative to the shallower-rooted Salix and Populus28.

Groundwater-dependent vegetation (GDV) polygons are Natural Communities Commonly Associated with Groundwater (NCCAG) vegetation polygons (n = 246,017 polygons). Vegetation polygons with an associated well (n = 3,113 polygons) are in blue, and those without (n = 242,904 polygons) are in grey.

a–h NDVI vs DTG plots (left) and ZDTG vs ZNDVI plots (right) for groundwater-dependent vegetation across California (a,b; n = 3,113 polygons with statewide CASGEM monitoring well data in grey/black); n = 26 polygons with Kern River Preserve monitoring well data in blue) and for three major groundwater-dependent vegetation genera: Salix spp. (c,d; n = 569 polygons), Populus spp. (e,f; n = 367 polygons) and Quercus spp. (g,h; n = 532 polygons).
To identify potential groundwater thresholds affecting groundwater-dependent vegetation, we selected vegetation polygons that exhibited a significant negative ZNDVI and ZDTG relationship (slope <0; p ≤ 0.05; n = 289 polygons; Supplementary Section 3). For each of the 289 vegetation polygons with a significant negative slope, we used a threshold of ZNDVI = −1 to interpolate the corresponding ZDTG value, where larger positive ZDTG scores correspond with deeper groundwater anomalies (Fig. 1). This approach provided a common metric to compute the local DTG level that resulted in a standardized reduction in greenness (that is, one standard deviation below the mean NDVI). For example, groundwater-related threshold responses were visible at two nature preserves along the Kern River and Santa Clara River (Fig. 5). At both sites, precipitous NDVI declines during drought years (Supplementary Fig. 1 and Supplementary Table 1) coincided with declines in DTG that lowered the water table and capillary fringe below the root zone for the dominant trees (that is, Populus fremontii and Salix laevigata; Fig. 5a,b,e,f, respectively). At both sites, a decline in NDVI due to vegetation crown die-back and mortality occurred in response to deeper groundwater levels during the 2012–2016 drought. The corresponding ZDTG thresholds (at ZNDVI = −1) along the Kern and Santa Clara rivers were 1.8 and 1.9 (Fig. 5d,h), respectively. However, hysteresis was observed in the ZNDVI versus ZDTG observations at the Santa Clara River site (Fig. 5h) due to an incomplete recovery in vegetation greenness despite a rebounded water table following the widespread mortality event that occurred there during the 2012–2015 drought period (and into 2016 along the Santa Clara River). A more complete NDVI recovery was observed at the Kern River site, because vegetation die-back was less severe as seen in the comparatively higher NDVI values during the drought. Drought-induced mortality along the Santa Clara River has previously been linked to rapid groundwater-level changes during the 2012–2016 drought12,13. Mortality and die-back of woody phreatophytes due to groundwater change and drought can result in cascading consequences such as loss of critical habitat for endangered and special status species and impairments to ecosystem structure and function27.

a–h, Site-specific NDVI and DTG trends at two nature preserves owned by Audubon and The Nature Conservancy, respectively, along the Kern River (Populus fremontii; gdeID: 134058, a–d) and the Santa Clara River (Salix laevigata, gdeID: 111026, e–h). a,e, NDVI time series for the entire 1985–2022 study period (grey line) and for the years where DTG data exist (coloured points corresponding to individual years). Drought years are shaded in gold. b,f, DTG time series at the vegetation polygon (DTG at veg) with the mean maximum rooting depth for the associated genus (solid horizontal black line) and the corresponding ZDTG threshold at the vegetation polygon (dashed horizontal red line in d and h, respectively). c,g, NDVI versus DTG (m) at the well associated with the vegetation polygon, where coloured points correspond to individual years (refer to a and e), and the black solid trend line is based on an ordinary least square linear fit. d,h, ZNDVI versus ZDTG with the ZDTG threshold set where the ordinary least square fit intersects ZNDVI = −1. Note that post-drought (after 2016) points for the Santa Clara River are considerably lower than the fitted lines in g and h, indicating a hysteresis response due to incomplete recovery of NDVI even after groundwater depth recovered to pre-drought levels.
The ZDTG thresholds determined at the Kern River and Santa Clara River sites are similar to the median ZDTG threshold of 1.8 for other vegetation across California (n = 289 polygons) that exhibit a significant negative correlation (slope <0; p ≤ 0.05) between ZNDVI and ZDTG (Fig. 6a). Statewide, the ZDTG threshold ranged between 1.3 and 2.5 (first and third quartiles), suggesting that most substantial reductions in greenness are most likely to occur when DTG falls 1.3 standard deviations below the mean. This 25th percentile threshold corresponds to a local baseline DTG of 4.0 m (Fig. 6b) and helps ensure a margin of safety when applying these findings to other sites and ecoregions. The 25th percentile local DTG threshold of 4.0 m is consistent with threshold findings from previous research on cottonwood (Populus spp.) and willow (Salix spp.) along the Santa Clara River12,13 and valley oak (Quercus lobata) along the Cosumnes River in California34. Whereas location-specific thresholds are ideal, these findings are helpful guidelines for other locations where groundwater management, modelling and scenario planning are needed but where major groundwater data gaps exist.

Threshold response histograms for groundwater-dependent vegetation (n = 289 polygons) exhibiting a significant negative (slope <0; p ≤ 0.05) ZNDVI and ZDTG relationship. a,b, Groundwater thresholds corresponding to ZNDVI ≤ −1 are reported as ZDTG thresholds (a) and DTG at vegetation (DTG at veg; in meters) (b). The 25th percentile (dashed line), median (solid line) and 75th percentile (dashed line) for ZDTG are 1.3, 1.8 and 2.5, respectively, and for DTG are 4.0 m, 7.9 m and 19.3 m, respectively.
Rooting depths are a good proxy for groundwater management
The use of ZNDVI and ZDTG regressions to quantify groundwater thresholds relies upon the occurrence of both groundwater depth threshold exceedances and corresponding ecological damage that are severe enough to be remotely sensed. To examine which ranges of groundwater depth support different levels of vegetation greenness, we compared the distributions of groundwater levels for vegetation that we classify as unhealthy (ZNDVI ≤ −1) and healthy (ZNDVI ≥ 1) vegetation versus baseline greenness (−1 < ZNDVI < 1; Fig. 7). Overall, DTG was deeper than published rooting depth values for the three dominant genera (Supplementary Table 2), except for Quercus, which has a reported average maximum rooting depth of 8.7 m across Quercus species. Our observation of DTG being deeper than published rooting depth values is consistent with our general understanding of water relations for phreatophytes, whose roots are preferentially distributed in aerated soils and the capillary fringe above the water table35 to avoid anoxic metabolic conditions and damage to roots. The capillary fringe receives water vertically from the water table through capillary rise, and the depth of the capillary fringe varies by soil type35. In coarse soils (for example, sand) the capillary fringe can be small and measured in centimetres, but in fine-grained soils, the capillary fringe can be >1 m in fine-grained soils35,36, which may account for the observed DTG values to be greater than documented plant rooting depths. For near baseline conditions (−1 < ZNDVI < 1), DTG (25th percentile) was within ~1 m of the maximum rooting depth, whereas groundwater levels were shallower for healthy vegetation and deeper for unhealthy vegetation. Differences in DTG between healthy and unhealthy vegetation was 0.9 m for all vegetation types but ranged between 0.4 and 1.4 m across the Salix, Populus and Quercus genera. The larger differences in DTG between healthy and unhealthy vegetation were observed for the shallower-rooted Salix and Populus, suggesting the importance of plant rooting depth relative to DTG. Thus, plant rooting depth information can be a good interim proxy for groundwater targets in the absence of complete groundwater data or in situ data, but more species-specific research on rooting depth, root structure and growth strategies are needed to inform local groundwater management.

a–l, DTG frequency distribution plots for all vegetation (a–c), Salix spp. (d–f), Populus spp. (g–i) and Quercus spp. (j–l) for unhealthy (ZNDVI ≤ −1; left column), near baseline (−1 < ZNDVI < 1; centre column) and healthy (ZNDVI ≥ 1; right column) vegetation. DTG at vegetation was calculated by subtracting the water surface elevation (NADV88) from the mean land surface elevation calculated at the associated vegetation polygon (Methods). The 25th percentile of DTG at vegetation locations for each subplot is indicated by a solid red, purple or blue horizontal line. The average maximum rooting depth for the associated vegetation in the subplots is indicated by the solid black horizontal line.
Anomalous NDVI patterns can locate potential drought refugia
Identifying local ecosystems that are resilient during drought is important for conservation planning and species recovery efforts; however, groundwater data are unavailable for 98.7% of the groundwater-dependent vegetation mapped throughout California (Fig. 3). To address this data gap, we utilized the 9.3 million ZNDVI data points across 246,017 vegetation polygons to map potential drought refugia (Fig. 8). These potential drought refugia are defined as places where vegetation is less likely to respond negatively to drought compared with the surrounding landscape and thus maintain critical habitat for associated species. Habitats supported by groundwater that can serve as drought refugia will become increasingly important amid climate change. Thus, identifying drought refugia can help practitioners to prioritize limited financial and natural resources to protect these critical habitats from groundwater depletion. To determine whether a groundwater-dependent vegetation polygon is a potential drought refugia site, we calculated the percentage of drought years (Supplementary Table 1 and Supplementary Fig. 1) where ZNDVI ≥ 0. Groundwater-dependent vegetation with a higher percentage of positive ZNDVI values during drought years are more likely to be drought refugia than vegetation with a lower percentage of positive ZNDVI values. On the basis of this analysis, only 1% (n = 2,483 polygons) of groundwater-dependent vegetation across California were identified as potential drought refugia, including Sequoia sempervirens within a protected redwood forest (Nisene Marks State Park) in Santa Cruz County, Populus fremontii adjacent to a wastewater treatment facility along the Mojave River and Populus balsamifera-Salix lasiolepis along a perennial stream reach in the Santa Clara River. Thus, drought refugia that are supported by shallow groundwater conditions can be influenced by both natural and anthropogenic factors.

a, Groundwater-dependent vegetation polygons are Natural Communities Commonly Associated with Groundwater (NCCAG) vegetation polygons (n = 246,017 polygons). Potential drought refugia are based on the frequency of ZNDVI ≥ 0 during drought years over the 1985–2022 study period. b, NDVI time series for select drought refugia examples, where drought years are shaded in gold.
Discussion
Our results highlight the utility of Z scores as a standardized metric and approach for identifying groundwater thresholds and targets for ecosystems across a diversity of site-specific conditions. For decades, determining groundwater thresholds and targets has been a localized process and has occurred primarily within jurisdictions where groundwater-dependent ecosystems are explicitly protected under legal groundwater-management frameworks. In many cases, findings from these limited local studies have not been directly transferable to other sites due to data gaps and cross-regional differences in local hydrology, vegetation and climate conditions. Combining large datasets with different temporal and spatial resolutions, such as satellite-based NDVI and ground-based well records, presents challenges as data must be curated to be relevant on similar timescales and to have ecologically appropriate scales. In this context, utilizing Z scores in ecosystem groundwater needs assessments provides a reproducible approach and is likely to be appealing to practitioners for its simplicity and robustness in acknowledging variability across local conditions. This is important so that ecosystem groundwater requirements can continue to be refined and assessed in California and other water-limited regions where groundwater-dependent ecosystems have been mapped.
Across California, we found that groundwater levels need to be maintained at levels 1.3 standard deviations below their local average (ZDTG < 1.3) to avoid a substantial, adverse ecosystem-level response; this ZDTG threshold corresponds proportionally to 4.0 m DTG across the range of local thresholds. Whereas variation in the ZDTG threshold exists across California and probably elsewhere, we found that groundwater levels for vegetation exhibiting baseline NDVI conditions were ~1 m to the rooting depth of most vegetation. Thus, taxonomically specific rooting depths inferred from community-level vegetation maps, which are more common to large management areas than long-term shallow well records, may be used as interim threshold proxies for impact analyses. This efficient detection method may support management planning not only for jurisdictions governed by sustainable groundwater-management legislation but also outside these regulated areas for minimizing environmental harm during project permitting and conservation efforts1,14.
Ensuring that water-limited ecosystems have access to groundwater requires an adaptive management approach with appropriate safeguards to operate within a sufficient margin to ensure that negative impacts do not cross tipping points into permanent ecosystem impairment14,20. This is especially important for GDEs that are defined by long-lived, foundation species such as riparian trees. Allowing or imposing severe perturbations, especially in systems with a high level of variability and uncertainty, may induce permanent ecosystem changes. Therefore, to effectively manage groundwater in these systems, there must be robust local groundwater-level data to evaluate impacts, set targets and adjust thresholds as needed. Currently, sparse groundwater data preclude the determination of localized thresholds and targets over broad areas. Nevertheless, long-term records of vegetation greenness, such as the 38-year Landsat NDVI dataset, which is ubiquitous and publicly available, can be leveraged along with climate data to compute ZNDVI and identify potential drought refugia. This approach supports a regional analysis that can inform where limited resources and policies should be prioritized so that groundwater can be protected for critical ecosystems in California and beyond.
Methods
Data acquisition
We accessed 38 years (1985–2022) of Landsat-derived normalized difference vegetation index (NDVI) data for groundwater-dependent vegetation from The Nature Conservancy’s GDE Pulse version 2.1 tool (https://gde.codefornature.org/, accessed 2 June 2023)37. The GDE Pulse tool provides average annual dry-season (1 July–30 September) data for groundwater-dependent ecosystems mapped across California from the California Department of Water Resources’ Natural Communities Commonly Associated with Groundwater version 2.0 dataset (https://www.scienceforconservation.org/products/natural-communities-groundwater-v2; n = 246,017 vegetation polygons)38; the resulting dataset comprises 9,343,646 annual NDVI data points in total across all polygons and years. NDVI is a common dimensionless index used in remote sensing to quantify vegetation ‘greenness’, which is calculated by subtracting the spectral values of the red band from the near infrared band and then dividing that value by the sum of both bands. NDVI ranges from −1 to 1, where values closer to 1 indicate vegetation with a higher density of green leaves, values of close to zero (<0.2) indicate bare ground or dead vegetation and values less than zero denote the presence of surface water39. Satellite imagery used in Fig. 2 are from the National Agriculture Imagery Program (NAIP; https://naip-usdaonline.hub.arcgis.com) and were accessed and mosaiced into annual mosaic composites for the 2012 and 2016 calendar years using geemap, which is a Python package for Google Earth Engine40.
NDVI and groundwater data were paired by linking each vegetation polygon to a nearby well. Each vegetation polygon was linked to a particular well if the vegetation polygon intersected a 250-m radius around the well. Our intent was to (1) increase the likelihood of selecting wells that monitor groundwater wells within shallow unconfined aquifers and (2) avoid the inclusion of wells from adjacent sub-watersheds. In addition, wells had to have at least three mean annual dry-season (1 July –30 September 30) groundwater-level observations within the study period (1985–2022). For polygons that met these criteria and had more than one nearby well, wells with the longest period of record and shallowest total well depth were selected. This resulted in only one well to be associated with each vegetation polygon (n = 3,128 well-polygon pairs). Groundwater data were accessed from the State of California (https://data.cnra.ca.gov/dataset/periodic-groundwater-level-measurements, accessed 2 June 2023). Because shallow groundwater observation monitoring well data are limited statewide (and for most places in the world), especially within riparian corridors and other groundwater-dependent ecosystems, we supplemented the California groundwater dataset with groundwater data from a dense shallow groundwater monitoring network along the Kern River at Audubon’s Kern River Preserve (2012–2022); these data were used to contextualize results presented in Figs. 4 and 5.
The depth-to-groundwater (DTG) values for the wells associated with each vegetation polygon were corrected for land surface elevation differences because monitoring wells are typically installed at higher elevations than groundwater-dependent vegetation, which commonly occupy low-lying areas near streams and wetlands. This approach assumes that water surface elevation is constant across a 250-m distance and corrects DTG for variations in topography. Local DTG at each vegetation polygon was calculated by subtracting the groundwater elevation in the associated monitoring well (measured above mean sea level in NAVD88) from the median land surface elevation of each vegetation polygon. Land surface elevations were determined from the US Geological Survey’s 1/3 arcsecond digital elevation model dataset (https://developers.google.com/earth-engine/datasets/catalog/USGS_3DEP_10m, accessed 6 April 2023).
Drought years within the 1985–2022 study period were determined using monthly standardized precipitation index data from https://www.drought.gov/historical-information, accessed 11 September 2023. The monthly standardized precipitation index data represent the percentage of California that was designated as each of the following categories: abnormally dry (D0), moderate drought (D1), severe drought (D2), extreme drought (D3), exceptional drought (D4), abnormally wet (W0), moderate wet (W1), severe wet (W2), extreme wet (W3), exceptional wet (W4) (Supplementary Fig. 1). Years where the average annual dry-season (July–September) monthly D0 classification, which is inclusive of the D0–D4 categories, was greater than or equal to 50% across the state were classified as a drought year (Supplementary Table 1). Note that this is a conservative definition of statewide drought, as local studies have indicated longer drought periods within different parts of California30.
Species-specific rooting depth data for groundwater-dependent vegetation in the Natural Communities Commonly Associated with Groundwater are from The Nature Conservancy’s Plant Rooting Depth Database (https://www.groundwaterresourcehub.org/where-we-work/california/plant-rooting-depth-database/, accessed 15 May 2023). Rooting depth data in this database were compiled from published scientific literature and expert opinion, and the original sources are cited therein. Genus-specific maximum rooting depth data were calculated by taking the average reported maximum rooting depth for all species within the same genus.
Statistical analyses
To calculate Z scores for NDVI (ZNDVI), we first calculated the mean and standard deviation for each vegetation polygon over the full 1985–2022 time series of annual dry seasons (July–September; n = 38 annual observations per polygon). Z scores were calculated for each annual observation at each vegetation polygon (n = 246,017 polygons) by subtracting the average annual dry-season NDVI observation (n = 38 annual observations per polygon) by the site-specific mean and then dividing by the standard deviation, resulting in 9,343,646 ZNDVI data points across California. Z scores for DTG (ZDTG) were calculated for each vegetation polygon with an associated well (n = 3,113 polygons) by subtracting the average annual dry-season DTG observation (n = 3–38 annual observations per polygon) by the site-specific mean and then dividing by the standard deviation, resulting in 36,381 ZDTG data points. The frequency (for example, continuous, annually, biannually, intermittent) of DTG observations was highly variable and contingent upon local monitoring efforts. Linear regressions between NDVI and DTG and ZNDVI and ZDTG were determined using an ordinary least-squares linear model (lm package in R).
Limitations
The groundwater-dependent vegetation from the Natural Communities Commonly Associated with Groundwater (NCCAG) dataset is a compilation of publicly available datasets, including the California Department of Fish and Wildlife’s Vegetation Classification and Mapping Program (VegCAMP), United States Forest Service’s Classification and Assessment with Landsat of Visible Ecological Groupings (CALVEG) and the United States Fish and Wildlife Service’s National Wetlands Inventory (NWI v. 2.0) datasets. These datasets were reviewed by experts at the California Department of Water Resources, California Department of Fish and Wildlife and The Nature Conservancy of California to map groundwater-dependent vegetation (phreatophytes) throughout California. Aerial imagery used in these vegetation mapping datasets had varying spatial and temporal resolutions, and it is possible that not all groundwater-dependent vegetation mapped in the resultant database are actually present for the entire study period. Additionally, these static delineated boundaries do not provide information on vegetation condition, and it is possible that ZNDVI changes or trends reflect land-use changes, afforestation/deforestation, wildfire events, flood scouring events or other fluvial morphological processes. Thus, local information on vegetation status may elucidate groundwater-related impacts. Despite the possibility of land-cover change influencing ZNDVI scores for individual polygons or local areas, using the longest period covered by the Landsat record is valuable for providing robust statistical moment values for mean and standard deviation that include many dry and wet years. In contrast, incomplete DTG well records are more likely to be prone to erroneous values of ZDTG, particularly if the available data do not represent the mean and standard deviation for the full 1985–2022 record due to skewed or insufficient data. Thus, shallow groundwater monitoring network improvements are essential to better characterize groundwater regimes and conditions within ecosystems, monitor resultant impacts caused by groundwater levels and mitigate and restore groundwater conditions within ecosystems to avoid harm.
Data availability
All data generated in this study have been deposited in Zenodo (https://doi.org/10.5281/zenodo.10627503)41. The raw data are publicly available and are accessible from the persistent web links provided in the Methods section and Supplementary Table 3.
Code availability
All code generated in this study have been deposited in Zenodo (https://doi.org/10.5281/zenodo.10627503)41. Data processing and analyses were performed using Python 3, and the figures were created using the programming language R (R Core Team, version 4.3.1) and Affinity Designer (https://affinity.serif.com/en-us/designer/).
References
Huggins, X. et al. Overlooked risks and opportunities in groundwatersheds of the world’s protected areas. Nat. Sustain. 6, 855–864 (2023).
Rohde, M. M., Froend, R. & Howard, J. A global synthesis of managing groundwater dependent ecosystems under sustainable groundwater policy. Groundwater 55, 293–301 (2017).
Saccò, M. et al. Groundwater is a hidden global keystone ecosystem. Glob. Chang. Biol. 30, e17066 (2023).
Jasechko, S., Seybold, H., Perrone, D., Fan, Y. & Kirchner, J. W. Widespread potential loss of streamflow into underlying aquifers across the USA. Nature 591, 391–395 (2021).
Graaf de, I. E. M., Gleeson, T., Beek van (Rens), L. P. H., Sutanudjaja, E. H. & Bierkens, M. F. P. Environmental flow limits to global groundwater pumping. Nature 574, 90–94 (2019).
Barlow, P. M. & Leake, S. A. Streamflow depletion by wells—understanding and managing the effects of groundwater pumping on streamflow. J. Hydrol. 352, 250–266 (2008).
Rohde, M. M. et al. A machine learning approach to predict groundwater levels in California reveals ecosystems at risk. Front. Earth Sci. 9, 784499 (2021).
Rodell, M. & Li, B. Changing intensity of hydroclimatic extreme events revealed by GRACE and GRACE-FO. Nat. Water 1, 241–248 (2023).
Liu, P.-W. et al. Groundwater depletion in California’s Central Valley accelerates during megadrought. Nat. Commun. 13, 7825 (2022).
Rohde, M. M. Floods and droughts are intensifying globally. Nat. Water 1, 226–227 (2023).
Condon, L. E., Atchley, A. L. & Maxwell, R. M. Evapotranspiration depletes groundwater under warming over the contiguous United States. Nat. Commun. 11, 873 (2020).
Williams, J. et al. Local groundwater decline exacerbates response of dryland riparian woodlands to climatic drought. Glob. Chang. Biol. 28, 6771–6788 (2022).
Kibler, C. L. et al. A brown wave of riparian woodland mortality following groundwater declines during the 2012–2019 California drought. Environ. Res. Lett. 16, 084030 (2021).
Saito, L. et al. Managing groundwater to ensure ecosystem function. Groundwater 59, 322–333 (2021).
Howard, J. K., Dooley, K., Brauman, K. A., Klausmeyer, K. R. & Rohde, M. M. Ecosystem services produced by groundwater dependent ecosystems: a framework and case study in California. Front. Water 5, 1115416 (2023).
Nelson, R. L. Water rights for groundwater environments as an enabling condition for adaptive water governance. Ecol. Soc. 27, art28 (2022).
Perrone, D. et al. Stakeholder integration predicts better outcomes from groundwater sustainability policy. Nat. Commun. 14, art3793 (2023).
Huang, F., Zhang, Y., Zhang, D. & Chen, X. Environmental groundwater depth for groundwater-dependent terrestrial ecosystems in arid/semiarid regions: a review. Int. J. Environ. Res. Public Health 16, 763 (2019).
Eamus, D., Froend, R., Loomes, R., Hose, G. & Murray, B. A functional methodology for determining the groundwater regime needed to maintain the health of groundwater-dependent vegetation. Aust. J. Bot. 54, 97–114 (2006).
Thomann, J. A., Werner, A. D. & Irvine, D. J. Developing adaptive management guidance for groundwater planning and development. J. Environ. Manage. 322, 116052 (2022).
Ellison, A. M. et al. Loss of foundation species: consequences for the structure and dynamics of forested ecosystems. Front. Ecol. Environ. 3, 479–486 (2005).
Froend, R. & Sommer, B. Phreatophytic vegetation response to climatic and abstraction-induced groundwater drawdown: examples of long-term spatial and temporal variability in community response. Ecol. Eng. 36, 1191–1200 (2010).
Keddy, P. A. & Reznicek, A. A. Great Lakes vegetation dynamics: the role of fluctuating water levels and buried seeds. J. Great Lakes Res. 12, 25–36 (1986).
Moore, D. R. J. & Keddy, P. A. Effects of a water-depth gradient on the germination of lakeshore plants. Can. J. Bot. 66, 548–552 (1988).
Sommer, B. & Froend, R. Phreatophytic vegetation responses to groundwater depth in a drying Mediterranean‐type landscape. J. Veg. Sci. 25, 1045–1055 (2014).
Scott, M. L., Shafroth, P. B. & Auble, G. T. Responses of riparian cottonwoods to alluvial water table declines. Environ. Manage. 23, 347–358 (1999).
Shafroth, P. B., Stromberg, J. C. & Patten, D. T. Woody riparian vegetation response to different alluvial water table regimes. West. N. Am. Nat. 60, 66–76 (2000).
Rohde, M. M., Stella, J. C., Roberts, D. A. & Singer, M. B. Groundwater dependence of riparian woodlands and the disrupting effect of anthropogenically altered streamflow. Proc. Natl Acad. Sci. USA 118, e2026453118 (2021).
Huntington, J. et al. Assessing the role of climate and resource management on groundwater dependent ecosystem changes in arid environments with the Landsat archive. Remote Sens. Environ. 185, 186–197 (2016).
Warter, M. M. et al. Drought onset and propagation into soil moisture and grassland vegetation responses during the 2012–2019 major drought in Southern California. Hydrol. Earth Syst. Sci. 25, 3713–3729 (2021).
Mahall, B. E., Tyler, C. M., Cole, E. S. & Mata, C. A comparative study of oak (Quercus, Fagaceae) seedling physiology during summer drought in southern California. Am. J. Bot. 96, 751–761 (2009).
Skiadaresis, G., Schwarz, J., Stahl, K. & Bauhus, J. Groundwater extraction reduces tree vitality, growth and xylem hydraulic capacity in Quercus robur during and after drought events. Sci. Rep. 11, 5149 (2021).
David, T. S. et al. Root functioning, tree water use and hydraulic redistribution in Quercus suber trees: a modeling approach based on root sap flow. For. Ecol. Manage. 307, 136–146 (2013).
Rohde, M. M., Sweet, S. B., Ulrich, C. & Howard, J. A transdisciplinary approach to characterize hydrological controls on groundwater-dependent ecosystem health. Front. Environ. Sci. 7, 175 (2019).
Eamus, D., Huete, A. & Yu, Q. Vegetation Dynamics: A Synthesis of Plant Ecophysiology, Remote Sensing and Modelling (Cambridge Univ. Press, 2015); https://doi.org/10.1017/CBO9781107286221
Lu, N. & Likos, W. J. Unsaturated Soil Mechanics (Wiley, 2004).
Klausmeyer, K. R. et al. GDE Pulse: Taking the Pulse of Groundwater Dependent Ecosystems with Satellite Data (The Nature Conservancy, 2019); https://gde.codefornature.org
Klausmeyer, K. et al. Mapping Indicators of Groundwater Dependent Ecosystems in California: Methods Report (The Nature Conservancy, 2018); https://www.groundwaterresourcehub.org/content/dam/tnc/nature/en/documents/groundwater-resource-hub/iGDE_data_paper_20180423.pdf
Roy, D. P. et al. Characterization of Landsat-7 to Landsat-8 reflective wavelength and normalized difference vegetation index continuity. Remote Sens. Environ. 185, 57–70 (2016).
Wu, Q. geemap: a Python package for interactive mapping with Google Earth Engine. J. Open Source Softw. 5, 2305 (2020).
Rohde, M. M. Data, code, and outputs for: establishing ecological thresholds and targets for groundwater management. Zenodo https://doi.org/10.5281/zenodo.10627503 (2024).
Acknowledgements
Financial support for this work came from the National Science Foundation (BCS-1660490 (J.C.S., M.B.S., D.A.R.), EAR-1700517 (J.C.S.), EAR-1700555 (M.B.S., K.K.C.)), the US Department of Defense’s Strategic Environmental Research and Development Program (RC18-1006 (M.B.S., J.C.S., D.A.R., K.K.C.), RC22-C1-3216 (M.M.R.)), the Bureau of Reclamation’s WaterSMART programme (R19AP00278 (C.M.A.)), an Ecosystem Technical Assistance grant issued by the Water Foundation and Audubon California (grant number 213012 (M.M.R.)) under a California Department of Water Resources Sustainable Groundwater Management Act Implementation Grant and the Zegar Family Foundation (award SB220237 (K.K.C.)). We thank R. Tollefson and staff at Audubon California for collecting and sharing groundwater data from the Kern River Preserve and to M. Stella for graphical assistance.
Author information
Authors and Affiliations
Contributions
M.M.R. and J.C.S. designed the research; M.M.R. developed the code; M.M.R. and J.C.S. analysed the data; M.M.R. wrote the paper; J.C.S., M.B.S., D.A.R., K.K.C. and C.M.A. contributed to the paper and provided intellectual support.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Peer review
Peer review information
Nature Water thanks Dylan Irvine and Christian Griebler for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Supplementary Information
Supplementary Sections 1–3.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Rohde, M.M., Stella, J.C., Singer, M.B. et al. Establishing ecological thresholds and targets for groundwater management. Nat Water 2, 312–323 (2024). https://doi.org/10.1038/s44221-024-00221-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s44221-024-00221-w
This article is cited by
-
Defining thresholds to protect groundwater-dependent vegetation
Nature Water (2024)
